THE SEA URCHIN, REVILED BY OCEAN REVELERS FOR ITS SPINES but beloved by sushi aficionados for its gonads, has a starring role in developmental biology.1 Much of what we know about the earliest stages of development comes from studies on sea urchins. Fertilization—the union of sperm nucleus with egg nucleus—was first observed in sea urchins. Sea urchins have also figured prominently in studies of what happens next: a series of cell divisions through which the fertilized egg, or zygote, is transformed into a ball of generic cells called a blastula. The cells in the blastula are generic in the sense that they have none of the distinguishing features of any of the cell types found in adult sea urchins, such as blood cells and neurons (sea urchins have neurons but no brains). These blastula cells are also generic in another sense: they generate all of the adult cell types. We call these generic blastula cells embryonic stem cells. This is as true for humans as for sea urchins.
The process whereby the zygote gives rise to the blastula, and the blastula gives rise to an animal with spines and gonads—or brains and gonads, as the case may be—is among the most wondrous in the universe. This process is also one of the most difficult for the human mind to digest. Our intuition, so useful in other scientific contexts, tends to fail us here, tends to lead us astray. There is also a lot at stake here. The ultimate goal, after all, is to understand how we come to be what we are, how you came to be you. It is not surprising, therefore, that this process, which we will simply call development, is a subject with a long history of dispute.
There are a number of points of contention, but for our purposes we can divide the disputants into two camps. In the first, we have those who assert that, despite appearances to the contrary, the zygote actually contained you. This is known as preformationism, which in its most extreme form asserts that your development was merely a matter of growth. The extreme form of preformationism is also the most primitive; far more sophisticated versions were developed during the eighteenth and nineteenth centuries.2 The more sophisticated versions of preformationism assert that you are latent in the zygote and your development is the process of the “latent you” becoming the “manifest you.” This latent you need not resemble the manifest you at all. But—and this is the essence of preformationism—the particular form that constitutes you is entirely present, however latently, in the zygote. Nothing in the environment contributes to your adult form. Nor did your manifest you come to exist as a result of development; it was there from the outset.
The last claim in particular distinguishes preformationism from the second type of developmental explanation, known as epigenesis.3 From the epigenesist perspective, you do not exist prior to development, either manifestly or latently. Rather, development is the process whereby you come to exist. Development is not just a matter of unfolding; it is a creative process. This is not to deny that the genes and other biochemicals in that zygote are essential for your development—they most certainly are. But they don’t contribute to your development by being a preformed you.
In the earliest (that is, seventeenth-and eighteenth-century) versions of preformationism, the preformed you was usually thought to reside in the unfertilized egg. Moreover, each egg was considered to be something like a set of Russian dolls, containing within itself the forms of all subsequent generations in increasingly diminutive sizes. Eve’s eggs contained within them all subsequent humans, including you. Though this seems absurd to us now, it was not, given the technology and knowledge existing at that time, a ridiculous hypothesis. Moreover, one of its considerable virtues was its ability to explain development without invoking any supernatural principles that violate the basic scientific principle of naturalism (as opposed to supernaturalism). Preformationists did not need any spooky stuff to explain how your mother’s egg became you. There is nothing mysterious about growth.
Advocates for epigenesis, however, claimed that despite its naturalism, preformationism was simplistic at best, and at worst downright wrong-headed. Epigenesists were generally more attentive than the preformationists to the complexities of development, but they had problems of their own. Most significantly, they could not account for the orderliness of this complex process—or how this ordered complexity could result from more simple, seemingly homogeneous conditions—without recourse to spooky stuff. For the preformationists, complexity was unproblematic; it existed from the outset, from the dawn of creation. Early epigenesists, however, like Darwin in a different context, had to explain how you get something very complex from something simple like a zygote. They could not find the explanatory principles they needed among the known physical laws, most notably Newton’s physics, so they posited an additional something that living things have and non-living things lack. The conceptions of this additional something varied greatly, but they had in common the notion that it is not material, and that it could only be demonstrated by pointing to the process itself. This does not count as a scientific explanation.
By the last few decades of the nineteenth century, experimental methods had improved enough to actually test some of the tenets of both camps. Two German scientists figured prominently in the debate at this point, Wilhelm Roux and Hans Driesch.
The Experiment and Its Aftermath
Driesch chose sea urchins for his study animal for the same reason that others before him had chosen sea urchins to study fertilization: their large, largely yokeless eggs. Sea urchin eggs are much larger than those of most other animals, including frogs and humans. This is especially important for monitoring not only fertilization but also the first few cell divisions, which occur within the confines of the zygote. As such, with each cell division, the cells become progressively smaller. But even after the first few cell divisions, the cells in the sea urchin embryo are relatively large, and hence easy to see with the microscopes available at that time, compared with cells in frog or human embryos at the same stage. The fact that sea urchin zygotes have very little yolk means that these cells are nearly transparent, which is also a plus.
A central issue in the debate at this time was, how do you get all the different cell types—blood cells, skin cells, cone cells, and so on—from an egg cell that doesn’t look anything like any of them? The state-of-the-art preformationist account in the 1890s, the one advocated by Roux, proposed that the chromosomes in the fertilized egg or zygote contain all of the determinants of the adult form.4 Parts of the chromosomes are then increasingly parceled out during each subsequent cell division, until all of the cell types have become fully differentiated. Whether the differentiated cell becomes a neuron, a muscle fiber, a blood cell, or whatever type depends on which particular bits of chromosome it contains. The idea appealed to many because it was straightforwardly mechanical.
Both Roux and Driesch set out to test this idea, but only Driesch succeeded in mastering the technical details.5 Driesch experimentally intervened right after fertilization, during those first few cell divisions. He managed to separate the cells in sea urchin embryos when they were only at the two-to eight-cell stages. To his surprise, each of the separated cells from the two-cell stage developed into a complete sea urchin larva, as did some from the four-and eight-cell embryos. According to Roux’s preformationist account, any cell isolated at the two-cell stage should have half of the chromosomal complement and could thus only become half of a sea urchin. As a result, Driesch’s sea urchin experiments amounted to a decisive refutation of Roux’s preformationism, and Driesch became an advocate for epigenesis.
Driesch reasonably concluded that in the very early embryo, each cell could somehow regulate its development so as to become a whole embryo. Processes such as Driesch envisioned for development, we now call self-organizing.6 Driesch then carried out another set of experiments, at much later stages of development, which were of even more significance. He ingeniously manipulated the position of cells such that those that would normally have become, say, spines, were moved to a position appropriate for mouthparts. According to Roux’s preformationism, the resulting embryo should have been a mess, with spines sticking out of its mouth. Instead, Driesch obtained normal embryos.
Driesch concluded that the potential of each cell is not determined by what chromosomes (genes) are in its nucleus, but rather on its position in the embryo. More generally, Driesch began to develop an epigenetic framework in which he anticipated the modern concept of gene regulation by the cellular environment. He proposed that cellular stuff outside of the nucleus (where the genes reside) influences what the nucleus (genetic material) does, which in turn influences what the cellular stuff does, and so on, in a reciprocal manner.7 This notion of reciprocal causation, or feedback—in which an action is both cause and effect—was new to science. It is now a fundamental principle in biology.
Like Roux, Driesch had commenced his experiments with the thoroughly naturalistic vision of explaining development using only the laws of physics and mathematics. As such, he found his results somewhat disillusioning. In fact, he became so overwhelmed by the complexity of development that he abandoned not only his simpleminded naturalism, but naturalism as a whole. He felt compelled to invoke a soul-like principle, which he called “entelechy” following Aristotle, to explain this complex process.8 He eventually abandoned biology for philosophy, to the detriment of both.
The Death and Resurrection of Preformationism
Driesch’s experiments doomed the existing incarnation of preformationism, but not preformationist thinking. In part, the reincarnation of preformationism reflected the inability of advocates for epigenesis to come up with a credible nonspooky mechanism. But there was another force behind the resurrection of preformationism: its intuitive appeal. The human mind seems predisposed toward preformationism; it taps into some naïve preferred modes of thought for scientists and nonscientists alike, when it comes to explaining complex phenomena like development (or evolution).
There are two related intuitions that undergird preformationism (as well as creationism). The first intuition concerns the “pre” in “preformationism”: the complexity must be there from the get-go, in the egg (or the mind of God). Call this the “complexity intuition.” The second intuition concerns the “form” in “preformationism”: the preexisting form in the egg (or the mind of God) directs the development of the adult form. Call this the “directorial intuition.”
A naturalized form of epigenesis must combat both intuitions without recourse to spooky stuff. Against the intuition that only complexity begets complexity, epigenesists must successfully argue that complexity can result from relatively simple initial conditions. Against the intuition that complexly ordered processes such as development require a central form-giving director, epigenesists must demonstrate that this order can result from local interactions at the cellular level, given certain initial conditions.9 This is not an easy task. Even Driesch, while anticipating the essential concepts of feedback, reciprocal causation, and self-organization, eventually copped out by resorting to the spooky notion of entelechy. Hence, many biologists, while acknowledging that Driesch’s experiments won the battle for epigenesis, could not bring themselves to concede that he had won the war. They felt vindicated in this regard by the rise of modern genetics.
Originally the science of genetics—as practiced by, for example, Morgan—developed completely independently of developmental biology. It was inevitable, however, that geneticists would eventually tackle development. They did so with a strong preformationist bent, largely unaware of the previous work by Driesch and others in developmental biology. On the new, genetically inspired preformationist account, your genes contain the complex form that is you, through which they direct the process of your development. Genetic preformationism was successfully packaged through a series of intuitively appealing metaphors. First came the metaphor of the “genetic blueprint,” then that of the “genetic recipe,” and finally the “genetic program.” Some combination of the recipe and program metaphors remains popular; they have in common the notion that the genes provide instructions, which cells execute. In genetic preformationism, the executive gene is scaled up to become the executive genome.10
These recipe/program metaphors are attractive because they connect the basic intuitions common to all versions of preformationism to human artifacts with which we are all familiar, from cakes to graduation ceremonies.11 Whatever their intuitive appeal, these metaphors cannot withstand even the most cursory scrutiny. You couldn’t cook up a single cell, much less a human being, given the instructions in the genetic recipe. Much of what you need to know lies elsewhere. More to the epigenesist point, most of the information in the recipe that goes into making you is not there from the outset. Rather, development is the process whereby this information comes to exist.12 The recipe is written during development, not prior to development.
The same goes for the metaphor of a genetic program. But the notion of a genetic program also suffers from another fatal flaw: the software-hardware distinction. The genes are supposed to comprise the software and the rest of the cellular constituents the hardware whose operation the genes instruct. But as we have seen throughout this book, our genes are as much part of our hardware as any other biochemicals, and as much instructed as instructors. In fact, the science of epigenetics makes sense only when genes are viewed as biochemical hardware.
From Stem Cells to Cone Cells
One of the most vexing problems for all versions of preformationism—from the most primitive (Eve’s eggs) to the most sophisticated (genetic programs)—is explaining how a ball of identical generic cells can give rise to specialized cells such as cone cells, neurons, muscle cells, and so forth. This process is called cellular differentiation. You will recall that Roux, the preformationist, thought differentiation was the result of the progressive partitioning of genes during development. The zygote has all of the genes, while cone cells retain only a small subset and muscle cells a different small subset. But we now know that all of the cells in our body are genetically identical.13 Cone cells have the same genes as liver cells, muscle cells, or any other cell type. What makes cone cells different from, say, heart muscle cells, is differences in gene expression, not differences in genes. These differences in gene expression are caused by epigenetic processes.
Think of the zygote in terms of a particular commodity: potential. Zygotes have lots of this potential; in fact, they have as much potential as it is possible to obtain. What sort of potential? The potential to become—through numerous intermediary states—any one of the more than two hundred cell types in the human body, plus the placenta. They are said to be totipotent, which basically translates as “total potential.” By the blastula stage (about 128 cells), the component cells have lost the capacity to become placental cells, but they can become any of the other two hundred plus cell types. These are the embryonic stem cells, which are said to be pluripotent. From the blastula stage on, some of that potential to become other cell types gets lost with each successive cell division. The daughter cells lose some of that potential in the mother cell; they become more limited in what they can—through numerous intermediary states—become. When that potential gets narrowed to a certain point, where the mother cells give rise to daughter cells that can only become a certain class of cell types—say, blood cells versus neural cells—we call them somatic stem cells. (Somatic stem cells are often misleadingly referred to as “adult stem cells” in the media.) Somatic stem cells have lost much of that potential possessed by the embryonic stem cells, but they still retain a large amount of potential, compared with cone cells or heart muscle cells. They are said to be multipotent, which means, roughly, the capacity to become multiple cell types—in contrast to both no potential (for example, cone cells) and almost total potential (as in embryonic stem cells). There are a number of types of somatic stem cells, including neural stem cells and blood stem cells.
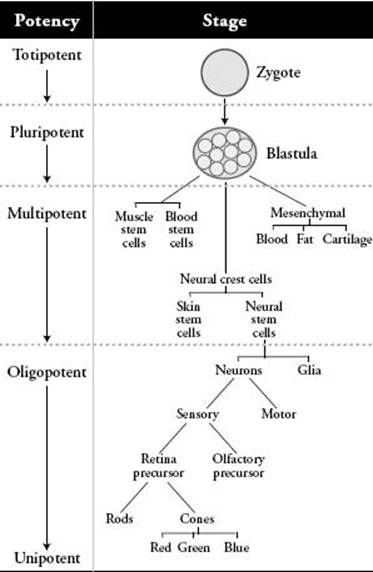
A schematic diagram of cellular differentiation from zygote to terminal differentiation of cone cells. Oligopotent cells have less potential than multipotent cells but more than unipotent cells. The branching within the oligopotent region illustrated here is highly speculative. Diagram by the author.
The transition from pluripotent embryonic stem cells to multipotent somatic stem cells, such as neural stem cells, is an epigenetic process during which an increasing number of genes are permanently inactivated (while other genes are newly activated). Differentiation continues past the multipotent somatic stem cell state—through further progressive epigenetic inactivation—until terminal differentiation into one of the two hundred plus cell types, such as cone cells or heart muscle cells. Cone cells and heart muscle cells are the end of the line differentiation-wise: cone cells can only beget other cone cells; heart muscle cells can only beget heart muscle cells. They have no more potential to become anything else.
A Third Form of Epigenetic Regulation
Both DNA methylation and histone modifications figure prominently in cellular differentiation, but there is another epigenetic mechanism at work here as well, which involves RNA.
We have already met one type of RNA involved in epigenetic regulation called Xist, which plays a central role in X-chromosome inactivation. Most of the RNAs with epigenetic functions are much smaller than Xist, however. Many, in fact, are extremely small. One important group of these tiny noncoding RNAs is called microRNAs.14
Epigenetic gene regulation by microRNAs works quite differently than either methylation or histone binding. Most significantly, microRNAs act later in the protein synthesis process. Recall from Chapter 2 that protein synthesis occurs in two stages. During the first stage, called transcription, messenger RNA (mRNA) is constructed from the DNA template. During the second stage, called translation, a protoprotein is constructed from the RNA template. Most epigenetic gene regulation occurs at the first stage, usually by inhibiting transcription. MicroRNAs, in contrast, exert their influence during the second stage, translation.
Though transcription is highly regulated, it is often the case that there are too many mRNA transcripts from a particular gene for a cell’s purposes. If a cell “decides” that this is the case, it deploys microRNAs to remedy the situation. The microRNAs identify the particular mRNAs that are overabundant and mark them for destruction. The microRNAs must physically bind to the messenger RNAs at a site where they are complementary. The microRNA doesn’t have to be perfectly complementary, just complementary enough to stick to the much larger mRNA. That means there can be many fewer types of microRNAs than there are types of mRNA transcripts. Sometimes the binding of a microRNA to an mRNA is sufficient to block the translation of the mRNA into protein. Sometimes the microRNA, in addition, attracts proteins, including enzymes, which actively degrade the mRNA. In either case, the net result is that there is less mRNA available to serve as a template for protein construction. This microRNA-based form of gene regulation, known as RNA interference, functions as a way to fine-tune the amount of protein that is made from a particular gene.15 Some have likened it to a light dimmer.
MicroRNAs play a major role in normal cellular differentiation.16 It appears that one of the primary functions of microRNAs at the cellular level is to stabilize the differentiation state of the cell. One study suggests that microRNAs don’t drive the differentiation but rather prevent dedifferentiation.17
Differentiation Is Reversible
You may have noticed that the process of differentiation superficially resembles Roux’s preformationist schema. Rather than a progressive partitioning of genes, there is a progressive partitioning of gene expression. But the resemblance is only superficial and the differences deep. The progressive partitioning of gene expression by epigenetic inactivation is not at all preformationist. Rather, as Driesch inferred, the fate of each cell, even well beyond the pluripotent stage, is largely determined by its position in the embryo and the nature of its neighbor cells with which it (chemically) interacts. These intercellular interactions influence the environment within the cell, which in turn influences which genes are epigenetically activated or inactivated. Hence, if you move cells around, especially during early stages of embryonic development, their fates change, just as in Driesch’s sea urchins, where cells normally destined for spinehood became part of the mouth when moved there.
The fact that differentiation is reversible also counts against any preformationist account. Under certain conditions, cells can dedifferentiate, that is, move back up toward “stemness.” This occurs naturally in injury repair, most spectacularly in amphibians, which can regenerate whole limbs and other organs.18 Dedifferentiation of skin, muscle, and bone cells is an essential part of this process. In mammals, dedifferentiation occurs in response to injury to cartilage and the peripheral nervous system.19 Some researchers seek to extend the range of dedifferentiation-based injury repair in humans based on amphibian models.20 But the most exciting potential for dedifferentiation therapies comes from stem cell research.
Stem cell biologists can now take a cell from the skin of an adult and, through biochemical subterfuge, transform it into the equivalent of an embryonic skin cell.21 Moreover, they can now take this skin cell–derived embryonic stem cell and cause it to redifferentiate into a neuron.22 Dedifferentiation is also one path to cancer, as we will see in the next chapter. But cancer cells themselves can be used to generate embryonic stem cells, in what is perhaps the most dramatic example of epigenesis.
Researchers were able to transplant human malignant melanoma cells into a chick embryo, a seemingly perverse procedure destined for failure.23 You would expect the melanoma cells, if they become integrated with the embryonic cells, to cause lots of problems. But in fact, the chick embryos were not compromised by the cancer cells. It’s not that the cancer cells died; they survived and divided at the normal rate. But the melanoma cells didn’t form tumors. Rather, the descendents of these cells actually assumed the identity of the normal cells from which the tumor cells were ultimately derived, in this case neural crest cells.24 They also migrated into the proper position for neural crest cells. It’s quite remarkable, really: you can use cancer cells to generate normal cells by exposing them to stem cells.
What happened to the cancer cells? Somehow, through interactions with the embryonic chick cells, the human cancer cells were epigenetically altered. These alterations caused the melanoma cells to dedifferentiate into stem cells, which then were able to redifferentiate into normal cells. This wasn’t the unfolding of a genetic script, so much as the writing (and rewriting) of it.
Epigenesis Without the Spooky Stuff
Epigenetics, it seems, provides the long-awaited naturalistic mechanism for epigenesis and should therefore spell the doom for preformationism. But the intuitive appeal of preformationism gives it a whack-a-mole capacity for resurrection. Every time the latest version gets crushed, a new version pops up to replace it. I will call the latest version of preformationism the “genetic-epigenetic program.”25 The genetic-epigenetic program metaphor acknowledges the central role of epigenetic events in development but views them through a preformationist lens. In essence, the idea is that the epigenetic events described earlier are programmed by the executive genome.
All of the problems with the “program” metaphor (as in “genetic programs”) apply to the notion of a genetic-epigenetic program—plus an additional one: in what sense are these epigenetic events programmed? Certainly not in the sense of “program” that most readers will bring to the table: a recipe-like set of instructions. As we have seen, the epigenetic changes in gene expression that determine a cell’s fate are largely determined by the position of that cell in the developing embryo. Therefore, it would be more apt to say that the genes are programmed by cellular interactions.
There are other, very minimalist senses of “program” that have become popular in the fields of artificial intelligence and artificial life.26 On this minimalist notion of program, a program provides a few basic rules, and the robots or cellular automata take it from there through interactions with their neighbors and the rest of their environment. But the idea of a central director is gone. This minimalist sense of program looks a lot like epigenesis. So the preformationist notion of an epigenetic (or genetic) program is either false or too squishy to be distinguished from epigenesis. In either case, we would be better served to drop the “program” metaphor altogether, and with it, the temptation to think of genes as software. Better to treat genes as concrete, material (biochemical) things, if we hope to understand the epigenetic processes that regulate them during differentiation.
A Note on the Stem Cell Controversy
Because of the protean nature of embryonic stem cells, they have the potential to revolutionize medicine. In principle you can place embryonic stem cells in any damaged part of the body, including the brain, and they will proliferate and differentiate into replacements for the damaged cells whatever they are. Heretofore untreatable conditions, such as injury to the spinal cord, can potentially be remedied in this way. Embryonic stem cells could help those who have been paralyzed for years to walk again—just one of countless potential applications.
Somatic stem cells can be utilized in some of the same ways. For example, neural stem cells might be able to do the job for spinal cord injuries. That would be a great boon because somatic stem cells are much easier to come by than embryonic stem cells. Until very recently, embryonic stem cells could only be harvested from blastula-stage embryos, a practice opposed by many on religious grounds. But we retain reservoirs of somatic stem cells even into adulthood, which is why somatic stem cells are often misguidedly referred to as adult stem cells (a misnomer since somatic stem cells are present in all but the earliest stages of embryonic development). It is therefore much easier to obtain somatic stem cells, and there is little opposition to this practice. But there are many applications for which embryonic stem cells remain more effective than somatic stem cells.27 This is why the recent studies demonstrating that stem cells can be induced artificially met with such fanfare. Here, potentially, is a way to generate more embryonic stem cells through dedifferentiation, one that won’t provoke the ire of opponents of embryonic stem cell research. But it will take years for scientists to convert this proof of principle into a viable technology for generating embryonic stem cells.28 In the meantime, embryonic stem cells harvested from embryos (blastulas) remain the best hope for many.
Opponents of embryonic stem cell research are motivated by a religious form of preformationism. They believe that a human soul is created when sperm meets egg. Secular versions of preformationism must also concede that the zygote is a human being, with all of the ethical implications that entails. From an epigenesist perspective, though, it makes no sense whatsoever to think of a zygote as a human being. A human being may or may not come to exist as a result of the processes initiated in the zygote. And zygotes produced by humans certainly have a much greater chance of becoming humans than those of, say, sea urchins; but that doesn’t make them actual humans. The same goes for the undifferentiated ball of cells that are harvested for embryonic stem cell research.
So at what point does the embryo become a human? Epigenesists can provide no hard and fast answers to that question. They can only note that human development is the process of becoming human¸ not the process of a preexisting latent human becoming a manifest human. Therefore, humanness is a matter of degree, not an absolute. It is up to us collectively as a society to decide how we should treat varying developmental degrees of humanness.
When Intuitions Lead Us Astray
From the outset, preformationism has had two considerable advantages over epigenesis: its intuitive appeal, and its naturalism. The drawback of preformationism has always been its failure in the experimental realm. Improved microscopes doomed the first (Eve’s eggs) version of preformationism. Driesch’s sea urchin experiments doomed Roux’s second-generation version. Despite these failures, preformationism remained viable because of its intuitive appeal, and because epigenesists like Driesch kept resorting to spooky stuff. When geneticists entered the fray, preformationism got a new boost, as genes seemed to provide an attractive preformationist mechanism, eventually expressed through two seductive metaphors—genetic recipes and genetic programs. Because it is purely metaphoric, this third-generation preformationism is inherently harder to put to the test. But epigenetics, properly construed, should spell the doom of preformationism.
The root of the term “epigenetic” is “epigenesis”—not, as is often supposed, “genetic.”29 Conrad Waddington, who coined the neologism in the 1940s, had in mind a naturalistic (nonspooky) form of epigenesis in which genes would play a prominent role.30 But the role that Waddington envisioned for genes much more closely resembled the executive cell view than that of the executive gene(ome).31 For Waddington, genes are as responsive to their cellular environment as their cellular environment is to them.32
Much has happened to the science of epigenetics since Waddington, and the temptation to view epigenetics through a preformationist lens remains powerful. The genetic/epigenetic program metaphor in particular is deployed effectively to this end. But modern epigenetics, as evidenced in the process of cellular differentiation, undermines preformationism as genetic program, most fundamentally, by challenging the view that genes are software that instructs the cellular machinery, viewed as hardware. Modern epigenetics makes sense only if genes are viewed as hardware, like other cellular constituents—as much instructed as instructor, as much directed as director, as much effect as cause.
Development does have a programmatic feel to it, but development is not programmed, in any but the squishiest sense of the term. Differentiation, for example, is an orderly process resulting from local cell-cell interactions. Whether a particular embryonic stem cell gives rise to cone cells or heart muscle cells depends on the history of these local interactions. Moreover, stem cells can induce dedifferentiation of differentiated cell types, not least in cancer cells. This dedifferentiation is often referred to as reprogramming.33 But here it is especially clear that the “reprogramming” isn’t done by the genes but to the genes. The same is true of the so-called programming of normal cellular differentiation. But all this talk of programming and reprogramming is a diversion from the concrete cellular interactions that are at the root of not only normal development but also cancer, which is the subject of the next chapter.