Daniel Amsterdam and Barbara E. Ostrov
This chapter describes biocides that serve as sterilants, disinfectants, and antiseptics, with emphasis on their use in health care facilities. Prevention of hospital-acquired infections is a national priority affirmed by the Institute for Healthcare Improvement (IHI) (1). These agents are key for prevention and are part of current standard infection control practices (2). An overview of the most widely used products, their active ingredients, their mechanism of action and spectrum of activity, and issues concerning resistance to these chemicals are addressed.
DEFINITIONS
In health care facilities, disinfectants are typically used in several different settings: by housekeeping to clean floors, walls, and other environmental surfaces; in laboratories, to decontaminate instruments and work surfaces; and in areas where instruments do not need sterilization but do require cleaning prior to reuse. As a group, these chemicals are referred to as instrument or environmental surface disinfectants. Antiseptics are used to reduce the microbial burden on skin or mucosal membranes.
Generally, a number of steps are required to properly achieve disinfection. These steps include cleaning, decontamination, low-level to high-level disinfection, and sterilization. Cleaning refers to the removal of organic material from an instrument or environmental surface. Decontamination is defined by the Occupational Safety and Health Administration (OSHA) as the removal, inactivation, and/or destruction of pathogens on a surface so they are no longer able to transmit infections and the surface is safe for handling, use, or disposal (3). Cleaning and decontamination are usually achieved by use of both enzymatic and nonenzymatic detergents and are primarily used to prepare devices or surfaces for subsequent disinfection or sterilization. Low- or intermediate-level disinfection is a process using chemical agents that will kill most pathogenic bacteria and fungi and nonenveloped viruses in a reasonable period of exposure time (≤10 minutes); typically, spores are not killed by these products. High-level disinfection is characterized by the product’s ability to kill or inactivate pathogenic bacteria and fungi and nonenveloped viruses as well as Mycobacterium tuberculosis; enveloped viruses; and, with prolonged exposure time, bacterial spores. The U.S. Food and Drug Administration (FDA) definition of high-level disinfection is an agent used for a defined contact time in order to achieve a −6 log10 reduction of Mycobacteriumspecies (4); several products on the market are capable of this level of disinfection. The primary active ingredients of such products belong to the chemical class aldehydes or oxidizing agents; however, some quaternary ammonium compounds and phenolics can serve as high-level disinfectants (Tables 12.1 and 12.2).


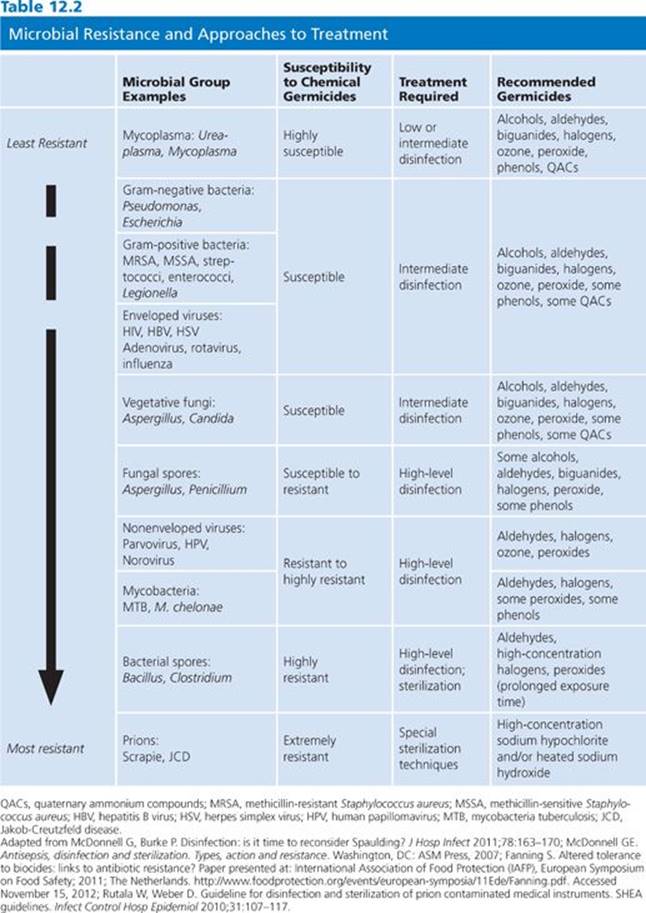
Sterilization processes must render surfaces and devices free of all living microorganisms, including spores. The sterility assurance level (SAL) defines the probability of survival of any viable organisms after sterilization treatment and is typically a −6 log10 or less than one in a million. Nonsterilization processes, although highly effective, produce instruments and surfaces that may yet contain some residual organisms and/or spores. It is important to appreciate that the use of chemical sterilants with shorter exposure periods will kill most microorganisms except large amounts of bacterial spores. Hence, sterilizing agents perform as high-level disinfectants in such situations. Unfortunately, some publications erroneously equate “disinfection” and “sterilization” or claim that items treated with short exposure times are “partially sterile.” The claim of chemical sterilization must be used to refer to absolute eradication of all types of microbes (4). Some agents that function as chemical sterilants include greater than or equal to 2.4% glutaraldehyde-based formulations, 0.95% glutaraldehyde with 1.64% phenol, 7.5% hydrogen peroxide, 7.35% hydrogen peroxide with 0.23% peracetic acid, 0.2% peracetic acid, and 0.08% peracetic acid with 1.0% hydrogen peroxide (see Table 12.2). These agents are most effective when cleaning precedes sterilization treatment and when standards for concentration, contact time, temperature, and pH are closely followed (4).
Antiseptics possess antimicrobial activity and may be used on living tissue to remove, inhibit the growth of, or inactivate microorganisms. The distinction between an antiseptic and a disinfectant is frequently ignored. However, the differences between a disinfectant and an antiseptic are important, and their applications are significantly different. A disinfectant is a chemical biocide used solely on inanimate objects or surfaces such as medical instruments or environmental surfaces; an antiseptic is to be used for living tissues. Some chemical agents, such as iodophors, can be active ingredients in either disinfectants or antiseptics. However, the precise formulations, patterns of use, and efficacy differ substantially. Consequently, products categorized as disinfectants should never be used as antiseptics and vice versa. Active ingredients used in antiseptics, including the iodine/iodophor group, biguanides (chlorhexidine), alcohols (usually ethanol or isopropanol), and phenolics (phenol and triclosan) have unique features, benefits, and drawbacks addressed in the remainder of this chapter.
SPAULDING CLASSIFICATION
The requirements for disinfection or sterilization of medical instruments, devices, or equipment can be more easily understood if these items are categorized based on the risk of infectivity involved with their use, as first suggested by Spaulding (5). The Spaulding classification system (Table 12.3) has been used since the 1950s by epidemiologists and microbiologists when discussing or planning strategies for disinfection and sterilization. The Centers for Disease Control and Prevention (CDC) has endorsed this system and has supported its use nationally and internationally (6).

Spaulding (5) defined three surface categories for treatment: critical, semicritical, and noncritical. Critical instruments or devices present the greatest risk of acquiring infection if the item is contaminated with microorganisms at the time of use. These are instruments or objects that are introduced directly into normally sterile areas of the human body. Examples include needles, scalpels, forceps, cardiac catheters, implants, and also the inner surface components of extracorporeal blood flow devices such as of the heart–lung oxygenator and the blood-side of hemodialyzers. Critical instruments must be sterilized by heat (such as steam autoclave or dry heat), ethylene oxide, hydrogen peroxide gas plasma, other low-temperature sterilization methods, and/or liquid sterilants between patient procedures.
Instruments or devices classified as “semicritical” have a lower risk of infection. Examples include flexible fiber-optic endoscopes, endotracheal tubes, bronchoscopes, laryngoscopes, respiratory therapy equipment, cystoscopes, vaginal specula, and urinary catheters. This category of devices comes in contact with mucous membranes and do not ordinarily penetrate body surfaces. Sterilization of many of these items, although desirable and often cost-effective if steam autoclaves can be used, is not absolutely essential. Semicritical instruments or devices should, at a minimum, be subjected to high-level disinfection using a product, usually a liquid chemical germicide, available for use as a sterilant but applied for a shorter exposure time so as to function as a high-level disinfectant. This approach has broad-spectrum efficacy and can destroy some bacterial spores, most fungal spores, all ordinary vegetative bacteria, mycobacteria, small or nonlipid viruses, and medium-sized or lipid viruses. In practice, good cleaning followed by high-level disinfection provides the medical practitioner assurance that these semicritical items are relatively free of pathogenic microorganisms.
“Noncritical” instruments or devices come into direct contact with the patient but usually only with intact skin. This category includes face masks, blood pressure cuffs, most diagnostic electrodes, and certain surfaces of X-ray machines. Depending on the particular item and the nature and degree of contamination during prior use, simple washing or scrubbing with a detergent and warm water may be sufficient to safely allow reuse. Transmission of infectious agents to patients from environmental surfaces typically involves a vector, commonly hospital personnel (7–10). Studies have indicated that skin surfaces harbor organisms such as Clostridium difficile (11–14), methicillin-resistant Staphylococcus aureus (MRSA) (7–9), and antibiotic-resistant Enterococcus strains. Controlling transmission of these pathogens can be accomplished by disinfection of environmental surfaces and use of proper handwashing with products employing an effective active ingredient. Thorough handwashing, along with the use of effective antiseptics, is one of the most important facets of the current overall infection control strategy (15,16). Beginning with the work of Semmelweis in the 1840s and confirmed by the World Health Organization (17), effective handwashing alone reduces the transmission of infectious agents. When successful handwashing is achieved and paired with effective biocide(s), transmission of microbes can be further reduced (18).
Housekeeping surfaces have very low potential for cross-contamination between health care personnel, patients, and medical equipment and/or instruments. Safe levels of microbes can be achieved by keeping these surfaces clean by using water and detergent or a hospital-grade disinfectant. A detergent designed for general housekeeping purposes (as indicated on the product label) would be adequate as well. In instances of spilled blood or a potentially infectious body fluid or culture specimen, an intermediate-level chemical disinfectant should be considered to render the environmental surface “safe.”
Spaulding’s original classification may be modified to include additional environmental surfaces, which carry the least risk of disease transmission (19). These additional environmental surfaces may contribute to secondary cross-contamination by the hands of health care workers or by contact with medical instruments that will subsequently come into contact with patients. Surfaces in this category encompass only those that come into direct contact with intact patient skin. These environmental surfaces may be further divided into (a) medical equipment surfaces such as adjustment knobs or handles on hemodialysis equipment, X-ray machines, instrument carts, or dental units, and (b) housekeeping surfaces such as floors, walls, tabletops, curtains between hospital beds, window sills, and so forth. As with noncritical medical instruments, adequate levels of safety for environmental surfaces may be achieved by simple cleaning with a detergent and warm water, cleaning with soap and water, or application of an intermediate- to low-level chemical germicide. Controlling the level of contaminant on environmental surfaces is usually accomplished with agents belonging to the quaternary ammonium and phenolic chemical classes.
In current hospital disinfection practice, antimicrobial wipes are used both to clean and to disinfect environmental surfaces. Some antimicrobial wipes remove a large proportion of microorganisms from environmental surfaces. However, germicidal activity associated with wipes might be limited due to brief application times and repeated use on multiple surfaces (20). Inadvertently, wipes may transfer microbial contaminants to other surfaces after multiple wipings (20). Studies assessing the effectiveness of germicide-containing wipes have revealed variable benefit as biocides (21). For example, the crucial eradication of C. difficile spores was established using hypochlorite-soaked wipes; peroxide-, biguanide-, and ammonium chloride–soaked wipes were not effective (21).
Additional factors may also influence the efficacy of the disinfection and sterilization processes. These include prior cleaning of the item; amount of organic and inorganic material present on the item; type and amount of microorganisms present on the item; exposure time to the germicide and the concentration of the germicide used; the physical features of the item to be treated, that is, the presence of difficult-to-reach cracks and crevices that need to be reached during treatment; the existence of biofilms; and the importance of temperature and pH of the disinfection or sterilization product and the item(s) to be treated (4).
Recently, the Spaulding classification has been reaffirmed to be as useful today as when first developed in 1957. However, this schema may be an oversimplification as it does not consider problems noted earlier nor issues such as complicated medical equipment that may require unique considerations, such as heat sensitivity, and it does not address the concerns about killing certain difficult-to-treat infectious organisms (e.g., prions) (see Table 12.2). Therefore, in some situations, selecting the best method of disinfection is challenging, even after evaluating Spaulding’s criteria. The efficacy and testing for the required levels of disinfectants and sterilants needs to constantly be reassessed in current times to address newly identified and changing pathogens as well as newer environmental surfaces or items that are used in the health care setting (4,22).
REGULATION AND TESTING OF PRODUCT CLAIMS
In the United States, disinfectants are regulated by the Environmental Protection Agency (EPA) and the FDA under the Federal Insecticide, Fungicide, and Rodenticide Act (FIFRA) (4). Some states have additional regulations. Under FIFRA, any product must be registered following approval of its claim of safety and effectiveness. The EPA regulates low- and intermediate-level disinfectants, and the FDA regulates high-level disinfectants. This division of regulatory responsibility parallels the use of the products, that is, the EPA controls environmental surfaces and the FDA controls medical devices. Antiseptics are regulated solely by the FDA as drugs. In the European Union (EU), in 2013, an updated directive regulating biocides was implemented, establishing a two-step process of evaluation at the EU level and product authorization at the member state level (23).
Regulatory Agencies
Testing regimens used for product claims are defined by the EPA and FDA in the United States. In the EU, requirements are defined by directives for biocides, disinfectants, and antiseptics. The European Committee for Standardization Technical Committee (CEN/TC) sets the standards for products in the EU. In the United States, test procedures used for meeting data requirements for disinfectants generally are those formulated by the Association of Official Analytical Chemists (AOAC), the American Society for Testing and Materials (ASTM), and the International Organization for Standardization (ISO). Internationally, regulatory agencies in some countries (e.g., Canada, Australia, and Brazil) use these or similar test methods. In the United States, historically, hospital disinfectants, whether used for noncritical; semicritical; or critical devices, were regulated by the EPA under the authority of FIFRA (4). Additionally, some regulatory agencies have adapted different end points that one must meet to make a claim. For instance, in the United States, the quantitative tuberculocidal test requires a 6 log reduction to make a claim for tuberculocidal activity, whereas in the EU, using the same procedure, a 5 log reduction is required. Hence, the same product marketed in the United States and the EU may have differing requirements needed to claim activity.
Testing Regimens
Test procedures have been established for disinfectants and antiseptics under the auspices of AOAC. The test protocols are primarily for determining the effectiveness of these agents against bacterial spores, fungi, vegetative bacteria, and mycobacteria, and they serve to determine accurate product claims. In addition, potency of ingredients must be verified periodically due to potential impact of continued use as well as mixing with organic material, such as blood or soil, on product efficacy (24). Various approaches are available, ranging from simple laboratory evaluation to assessment under actual clinical conditions. These approaches allow not only for evaluation of biocide activity but also new product development, regulatory approval, labeling of products, and standardization. Efficacy claims as reported in the literature have validity under described test conditions. However, one must keep in mind the potential impact of environmental and institutional variability when generalizing these results to one’s local hospital practice.
The effectiveness of a disinfectant or antiseptic is assessed by testing in three stages. The first stage verifies whether a chemical has adequate antimicrobial activity; suspension tests are typically used (Table 12.4). The second stage is performed using methods that simulate real-life conditions. Ingredient activity with testing using standardized conditions, such as duration of contact time and temperature, are determined (e.g., surface testing). The third phase is “real life” or in-use tests. In-use tests also verify disinfectant effectiveness after employment for a typical period of time. These tests are discussed in detail in the following text.


Laboratory test methods include determination of chemical uptake into cells, lysis of microorganisms and release of intracellular contents, alteration of cell wall permeability, alterations of cell membrane and metabolic function, enzymatic activity, disruption of biochemical pathways, and observation of microbial cell changes via microscopic visualization (25). Antimicrobial activity of the active ingredients is measured by determining the minimum inhibitory concentration (MIC), lowest concentration of the biocide that inhibits the growth of the microorganism, and minimal bactericidal concentration (MBC), the lowest concentration of the germicide that kills the organism. These test assays are modeled after those used for evaluating antimicrobial activity (see Chapter 3). Table 12.5 lists the MIC for several common microbes and germicide ingredients (26). Investigators often express the effectiveness of disinfectants in terms of decimal reduction time, or D-value, which is a commonly used measure of product efficacy. The D-value represents the time it takes, while at a constant temperature, to measure a predetermined significant reduction, usually 90% or greater, of the microbial load following treatment with a biocide (27). The microbicidal effect (ME) can also be determined for a product by subtracting the log number of microbes before and after treatment. The ME of 1 means there was 90% reduction of initial bacteria numbers, and ME of 2 equates to 99% killing. It is generally preferred that the ME is greater than or equal to 5, or that at least 99.999% microbes have been killed (24).

Ideally, a test protocol should simulate the conditions for which the product is intended. In vivo systems, involving human tissues or equivalent experimental models, and in situ systems, testing in real conditions, should be used when assessing antiseptics and disinfectants. For measuring in vivo or in situ activity, there are several protocols used to test formulations and preparations (see Table 12.4).
Suspension testing is a commonly used technique for assessment of efficacy of biocides (28). In this method, the test microbes are exposed to a series of chemical germicide dilutions and the MIC and/or MBC is determined. This method may be limited due to germicide interactions with organic or inorganic constituents in the growth media (e.g., halogens and aldehydes) or chemical agent interactions with the surface (e.g., chlorhexidine with certain fibers) (28). Analysis of the exposure time and the concentration of the biocide, along with other physical factors such as pH, temperature, and interfering substances (examples are organic materials such as serum or blood and inorganic materials such as hard water and soil), are crucial to determine the MBC and are practical considerations for use of the products in hospital settings.
Surface testing is necessary to assess the efficacy of the germicide on the surfaces for which they will be used. This is important for products to be used in hospital surfaces, including antiseptics, disinfectants, and sterilants. Typical methods used are carrier tests, simulated-use tests, and in-use testing. In the United States, stainless steel and porcelain surface items are used to perform carrier testing of products. The microbes are placed onto the item, with or without interfering substances such as soil, and then exposed to the germicide product. The test microbes are then assessed for survival via standard culture techniques. Simulated-use tests apply a sample inoculum of microorganisms to a surface to mimic actual use of the item and then wipe the surface with germicide. Recovery and culture then determines the survivability of the microbe. In-use testing is similar except that the assessment is performed in actual situations and conditions. This testing method may be recommended to evaluate the success of a product over time when used in hospital settings. The germicides must be assessed in-use with the most resistant organisms, such as spores, (“worst-case scenario”) to ensure actual sterilization is achieved. Detailed test methods, guidelines, and standards can be reviewed in McDonnell (28).
In-use testing for antiseptics requires in vivo assessment of products. Comparing in vivo studies is complicated because methods used vary greatly. For example, when studying different alcohol-based waterless antiseptic handrubs, methodologies vary as to (a) whether or not hands are purposely contaminated, (b) the volume of test substance used, (c) the time period the solution is in contact with the skin, (d) the methods used to recover organisms, and (e) the method of expressing efficacy. Several protocols have been used for determining the effectiveness of such antiseptics (29). Recent approaches have included a two-step process of in vitro testing followed by in vivo treatment evaluation (30). Alternatively, sequential in vivo application of two different antiseptics has been found to be highly effective (31). Product efficacy can be determined in vivo by using volunteers. In the Vienna test model, a variant of in-use testing, for example, a product is compared to standard disinfection assessed during parallel use with the same volunteers (32). This standardized approach allows for comparison of efficacy, by using experimental contamination of volunteers, and assessment of the survivability of the bacteria before and after disinfection. An exhaustive review of additional methodologic designs is provided by Hobson and Bolsen (33).
Numerous concerns about the traditional approaches to product testing have been posited (34–36). These concerns include lack of standardization of inocula, organisms’ resistance patterns, variations of the porcelain carriers used in the sporicidal tests, variations in the number of times the carriers are reused and processed, and uncertain relationship between the tested carrier surfaces and the surfaces to be disinfected in real-life situations. In response to these criticisms, alternative procedures have been proposed and adopted. The EPA adopted the use of a quantitative suspension test for measuring tuberculocidal activity of disinfectants (37) and modification of this methodology was adopted by ASTM (29). Further refinements have been developed to improve test reliability claims against mycobacteria (38). The updated quantitative carrier test (QCT) method has the following advantages: fully quantitative; minimizes the potential for false positives; measures kill under ideal and stringent conditions; increases the reliability of the results; can be applied to a wide variety of microorganisms, vegetative and spore forming; and eliminates the potential loss of organisms from wash-off during the exposure period. The QCT encompasses two separate parts, QCT-1 and QCT-2, which differ in the hard surface used (glass vs. stainless steel) and the type of organic soil challenge incorporated into the test. QCT-1 tests the disinfectant under more ideal conditions, using a glass surface, and without including organic soil, while QCT-2 tests the disinfectant under less ideal conditions. The latter is achieved by use of stainless steel (a surface that can have more variability), less disinfectant, and an organic challenge added to the suspension. QCT-1 has been validated for use with vegetative bacteria, including mycobacteria, bacterial spores, and fungi, whereas QCT-2 has been validated for use with viruses and protozoans in addition to vegetative bacteria, mycobacteria, bacterial spores, and fungi. The data obtained from these procedures allows one to calculate a log reduction value for the potency of the germicide. Each regulatory agency may choose to set their end point, the SAL, for determining product efficacy.
EPA developed a method for evaluating chemical germicides for virucidal activity, a method also accepted by the FDA for those products under its authority. Claims made based on testing against specific viruses, such as norovirus and hepatitis B and hepatitis C viruses, have been allowed based on testing using the surrogate viruses feline calicivirus, duck hepatitis virus, and bovine diarrhea virus, respectively (39).
Although some chemical germicides maybe labeled as sterilants, guidelines from the CDC, FDA, Association for Professionals in Infection Control and Epidemiology (APIC), and other professional organizations do not recommend the use of these germicides for sterilization unless there are no other available products. Unlike terminal sterilization processes (such as steam, ethylene oxide, and peroxide gas plasma), where the items can be maintained in a microorganism-free state by packaging after sterilization, items so treated using liquid processes cannot be maintained in a sterile state when they are removed from the solution. Sterilization claims can be obtained for the liquid processed products using the AOAC sporicidal test (40). Humphreys’ (41) report identifies the currently available methodologies and standards for sporicidal products.
ACTIVE INGREDIENTS
The active ingredients in antiseptics and disinfectants achieve their effects through interactions with the microorganism cell surface followed by penetration into the cytoplasm and action on cellular targets. The variable response to these products may be due to composition of the cell surface, change in the environment, and increasingly identified resistance to germicides. The following sections describe mechanisms of action and indications for commonly used products (summarized in Table 12.1; Fig. 12.1) (25,42).

BIGUANIDES
Chlorhexidine
Biguanides are compounds that contain the C2H5N7 component. Chlorhexidine, a substituted biguanide, has a high degree of antimicrobial activity, low mammalian toxicity, and the ability to bind to the stratum corneum layer of the skin and to mucous membranes (43). The bactericidal activity of chlorhexidine is more potent than that of monomeric biguanides, hence is the only biguanide discussed in detail in the following text (44). These unique characteristics make it particularly attractive as an active ingredient in antimicrobial skin preparations. Chlorhexidine’s general chemical structure is shown in Table 12.1.
Practically insoluble in water, chlorhexidine reacts with acids to form salts with varying degrees of solubility. Recognized as an effective antimicrobial, chlorhexidine is the active ingredient in a number of products: preoperative skin preparations, surgical hand scrubs, health care personnel handwash products, skin cleansers, acne creams, oral products (such as mouthwashes), burn ointments, and wound protectants; it has also been incorporated into products as a preservative.
Mechanism of Action
The activity of chlorhexidine relates to the interaction of the germicide with the cell surface (see Table 12.1 and Fig. 12.1), causing changes in the lipids in the cell membrane (45–47). Changes in the integrity of the membrane result in loss of membrane function. Experiments using 14C-chlorhexidine indicate that chlorhexidine rapidly enters the cells of both bacteria and yeasts (48,49). The bacterial cell wall is generally negatively charged and is rapidly neutralized in the presence of chlorhexidine (43), indicating the reaction of chlorhexidine with the cell membrane. The outer membrane structure in gram-negative bacteria is stabilized by the interaction of divalent cations and the negative charges associated with lipid A of the lipopolysaccharide (LPS) in the cell membrane. It is hypothesized that polycationic molecules, such as chlorhexidine, can promote their own uptake by displacement of divalent cations associated with the LPS. Alterations in the LPS can lead to changes in susceptibility to antimicrobials (50), whereas disruption of the cell membrane results in leakage of cellular material from the cell. The leakage in Escherichia coliand S. aureus is biphasic, depending on chlorhexidine concentration (47).
At low concentrations (up to 200 μg/mL), chlorhexidine inhibits membrane enzymes and promotes leakage of cellular constituents. Cells treated with bacteriostatic concentrations lost up to 50% of their K+but can recover upon neutralization of excess chlorhexidine (43,51). At higher concentrations, cytoplasmic contents are coagulated (52) and cells begin to leak higher molecular weight molecules such as nucleotides (43). Uptake of bis-(1,3-dibutylbarturic acid) trimethane oxonal and propidium iodide, which reflect membrane potential and membrane integrity, respectively, was directly related to chlorhexidine concentrations (0.003 to 0.3 mmol−1), indicating changes in membrane structure with increasing concentrations of chlorhexidine (53).
The effect of chlorhexidine on mycobacterial cells is bacteriostatic, not bactericidal. The treatment of mycobacterial cells with ethambutol (an antituberculosis drug known to inhibit the synthesis of specific components of the mycobacterial cell wall) potentiates the activity of chlorhexidine against Mycobacterium spp, indicating that chlorhexidine needs access through the cell membrane to exert its activity (54).
For yeast, chlorhexidine can similarly damage the cytoplasmic membrane. Treatment of Saccharomyces cerevisiae cells with chlorhexidine resulted in the release of pentose indicating increased oxidative stress and cellular leakage from these organisms (55). In Candida albicans, sublethal concentrations of chlorhexidine resulted in loss of cytoplasmic components and coagulation of nucleoproteins (56) and prevented the outgrowth but not the germination of spores (57).
The general mechanisms of chlorhexidine can be summarized as follows:
■ Damage to cell membrane by interaction with lipid layer
■ Leakage of cellular constituents at low-level exposure
■ Precipitation of nucleic acids and proteins at high-level exposure
■ Coagulation of intracellular and disruption of enzymatic pathways at high-level exposure
Spectrum of Activity
A comprehensive review of in vivo and in vitro effectiveness of chlorhexidine activity has been reported (43). MICs for bacteria and fungi are presented in Tables 12.6 and 12.7, respectively. These data indicate the broad spectrum of activity of this ingredient, which is effective against gram-positive and gram-negative bacteria, yeasts, and molds.



Studies of a 0.05% chlorhexidine solution confirm the rapid inactivation of gram-positive and gram-negative bacteria, yeasts, and molds (43). Following a 10-minute exposure to this solution at 18°C to 21°C, only resistant microorganisms, such as Pseudomonas stuartii, Streptococcus faecalis, Bacillus subtilis, and C. difficile, did not show substantial reduction. The MIC values for B. subtilis and Clostridium spp(see Table 12.6) are relatively low, indicating the bacteriostatic effect on these organisms. All of the other bacterial species tested showed a greater than 2 log10 reduction in 10 minutes. Gram-negative bacteria were generally more susceptible (4.0 to 6.7 log10reduction) than gram-positive bacteria (2.1 to 5.8 log10 reduction) under the conditions tested (58). Treatment of E. coli and Klebsiella pneumoniae with sublethal concentrations of chlorhexidine resulted in lower pathogenicity of these organisms when injected into mice (58). With the exception of the more resistant organisms listed earlier, among the gram-positive bacteria, the cocci (2.1 to 5.8 log10 reduction) were generally more resistant than the gram-negative bacilli (3.6 to 4.8 log10 reduction).
Data indicate that chlorhexidine may be capable of inhibiting spore growth and germination but that it does not kill the spores. As shown in Table 12.6, the MIC for spores is quite low. Data indicate that both B. subtilis and C. difficile spores show less than 1 log10 reduction when exposed to a 0.05% solution of chlorhexidine. Sporicidal activity can be achieved with chlorhexidine at higher temperatures (57).
Chlorhexidine has antiviral effects due to its interaction with viral membrane lipids. Viruses can be grouped into those with an outer lipid membrane composed of lipoproteins and glycoproteins around the protein coat (enveloped viruses) and those that are nonenveloped (i.e., lack a lipid membrane). Due to its solubility characteristics, chlorhexidine is more active against enveloped viruses with a lipid membrane (Table 12.8), such as HIV. Studies confirm the effectiveness of chlorhexidine against HIV. Preparations containing chlorhexidine (a 4% and a 0.5% solution in 70% alcohol) were completely effective against HIV-1 after 15 seconds of contact. An aqueous solution of 0.05% was equally effective in 1 minute (59). Aqueous chlorhexidine gluconate at 0.2% was effective but at 0.02% was not effective (60).

In vivo studies using chlorhexidine formulations provide further evidence of the activity of this germicide. The chemical components and the solubility of various chlorhexidine salts can make a difference in the activity of the formulation. When used in the presence of organic ions such as soaps, sodium lauryl sulfate, sodium carboxymethyl cellulose, alginates, and some dyes, chlorhexidine may be less effective. Many of these compounds are commonly used in formulations for antiseptic preparations. Although there may be no visual evidence of incompatibility, the chlorhexidine can be incorporated into micelles and not be readily available to interact with microorganisms (43). The addition of anionic thickeners or emulsifiers also serves to inactivate the long-lasting antibacterial activity (persistence) of chlorhexidine (61).
Chlorhexidine has been incorporated into detergent and alcohol preparations, powders, mouthwashes, and polymeric preparations for a variety of uses. Given its ability to bind to the stratum corneum layer of the epidermis, chlorhexidine exhibits persistence and substantivity; that is, it exerts its effect long after it has been used, and the more it is used, the greater the reduction in the resident flora of the skin. This activity is especially relevant for its use as a surgical scrub and in surgical site preparation.
As a surgical scrub, chlorhexidine has been prepared as 4% and 2% aqueous solutions as well as alcohol-based formulations of different concentrations. Chlorhexidine is effective in removing transient microflora from artificially contaminated hands (62). The effects are immediate and long-term (persistent or residual) on resident microflora (63). A study examining the difference between aqueous and alcohol preparations found that a formulation containing 1% chlorhexidine in 61% ethanol resulted in significantly greater microbial reduction than a 4% aqueous solution or the 61% ethanol vehicle alone (64). When used as a health care personnel handwash, chlorhexidine has typically been employed as the active ingredient in aqueous or alcohol-based solutions. Comparison of four chlorhexidine formulations (two 4% aqueous formulations, a 2% aqueous chlorhexidine formulation, and an alcoholic 0.5% chlorhexidine formulation) versus a nonantiseptic soap revealed that after the initial wash with each, there was no difference in microbial reductions. However, as more washes were performed, there was a significant difference between the four chlorhexidine-containing formulations as compared to the soap, although there was no difference between the chlorhexidine formulations (65).
Incorporation of chlorhexidine into surgical skin prep (2% chlorhexidine in 70% isopropyl alcohol) has proven to be effective. The chlorhexidine solution significantly reduced abdominal and inguinal microbial counts from baseline for up to 24 hours. It was more persistent in the abdominal region than were two other preparations containing isopropyl alcohol or chlorhexidine alone (66).
Chlorhexidine-containing powders used in routine umbilical decontamination in a neonatal unit controlled MRSA (67). Several studies have been done comparing infection rates when chlorhexidine gluconate and povidone-iodine were used as skin preparations in patients with central venous catheters. Comparison of catheter-related bloodstream infections indicates that chlorhexidine gluconate was superior; however, all the studies were limited by the small numbers of subjects (68–70). A meta-analysis indicates that chlorhexidine reduced the risk of catheter-related bloodstream infections by 50% (71). Several studies have assessed whether catheters impregnated with chlorhexidine and silver sulfadiazine (CHX-SS) reduced catheter-related bloodstream infections. When comparing patients with short-term indwelling catheters (median duration of 6 days) and those with long-term indwelling catheters (median duration of 12 days), a meta-analysis indicated that those in the short-duration group using CHX-SS catheters had a significant reduction in the incidence of catheter-related bloodstream infections, whereas the longer duration use group showed no difference between the CHX-SS catheters and standard catheters (72). In vitro studies with CHX-SS supported the data from the in vivo studies (73).
The incorporation of chlorhexidine into oral solutions goes back over 30 years (74), including mouthwash, varnish, and slow-release dental treatments. Chlorhexidine in varnish used on orthodontic patients helps prevent caries. Newer techniques such as epifluorescence microscopy can verify the dental effectiveness of chlorhexidine and other antimicrobial ingredients (75). Data indicate that there is a significant reduction in the level of Streptococcus mutans 1 week after the application of a sustained-release varnish. The decrease in S. mutans was measured for up to 3 weeks after the application. However, chlorhexidine-coated toothbrushes used for 30 days did not exhibit less bacterial contamination than untreated toothbrushes (76). A recent study compared the efficacy of chlorhexidine and a garlic extract in reducing colony-forming units of S. mutans. Although the chlorhexidine was beneficial, the garlic extract was more effective. It seems unlikely that the general public would be willing to perform regular garlic extract rinses, however (77). Studies have also shown high activity against typical oral flora when exposed for 1 minute to chlorhexidine digluconate 0.01% to 0.5% and no evidence of significant antimicrobial resistance was detected (78). Chlorhexidine may reduce ventilator-associated pneumonia (79) and possibly postdental implant bacteremia (80). Even with this known effect, current literature does not support a role for chlorhexidine rinses to prevent endocarditis in high-risk patients but instead recommend consistent comprehensive oral hygiene (81).
HALOGEN RELEASERS
Halogen releasers function as oxidizing agents, with bromine, chlorine, and iodine commonly used as disinfectants and antiseptics. Only certain halogen-containing species with iodine and chlorine are active antimicrobials due to their solvency and chemistry. Only these halogen releasers will be reviewed in this section.
Iodine and Iodophors
Iodine in the form of a tincture (alcohol extract of a low volatility substance) has been used since the early 1900s as a preoperative skin preparation. Cutaneous toxicity limited the use of iodine, whether in an aqueous or tincture preparation. The use of iodophors, which are complexes of iodine with a carrier such as polyvinylpyrrolidone (PVP) (see Table 12.1), allows for the slow release of free iodine, the active species, thereby reducing the preparation’s toxicity and staining without limiting the antimicrobial activity. Iodophors have been used as both antiseptics and disinfectants in the form of solutions, ointments, and aerosols. Iodophors are classified as low- or intermediate-level disinfectants (82) because they are not sufficiently sporicidal in an appropriate short application time. The amount of free iodine present in iodophors is difficult to determine. Concentrated solutions contain less free iodine than those that are diluted (83), and free iodine is the active species, whereas the PVP or other molecules with which the iodine is complexed act as carriers. A comparison of several commercial formulations of 10% polyvinylpyrrolidone iodine (PVP-I) had two orders of magnitude range of free iodine, hence a wide range of antimicrobial activity (84). Improperly diluted ionophore formulations allow for less free iodine and lower antimicrobial effects (85). This was the presumed cause of the outbreak of bacteremia associated with Pseudomonas aeruginosa when using a contaminated 10% PVP-I solution (86). Today, the primary use of iodophors is as an antiseptic for surgical scrubs and skin preparations.
Mechanism of Action
The killing effect of iodine and iodophors is related to the concentration of the free molecular iodine, whose concentration correlates with antimicrobial activity (83,87). The exact mechanism of action of iodine is not known. Its ability to penetrate the cell wall rapidly may be the primary mode by which it exerts its antimicrobial activity (see Table 12.1 and Fig. 12.1). Iodine can cause disruption of cells or cellular components as follows:
■ Reactions with N–H groups of amino acids during which N-iodo compounds are formed, resulting in destruction of proteins
■ Irreversible oxidation of -SH groups of cysteine, leading to loss of protein disulfide bonds (84,88)
■ Iodination of phenolic and imidazolic groups of the amino acids tyrosine and histidine and iodination of the pyrimidine derivatives of cytosine and uracil, leading to steric hindrances in hydrogen bonds and denaturation of DNA
■ Iodine binding to unsaturated fatty acids, altering the physical properties of lipids and hence lipid-containing membranes
For a more detailed description of the biochemical properties of these agents, the reader is referred to Gottardi (84).
Chlorine
The antiseptic- and disinfectant-oxidized chlorine compounds are formed in water. These include elemental chlorine, hypochlorous acid, and hypochlorite ion; the latter being the most widely used form. The antimicrobial action of available chlorine depends on the concentration, temperature, and pH of the solution; higher pH solutions have diminished antimicrobial activity. Many chlorine products have an irritating and corrosive nature, limiting their clinical use.
Disinfection of water is a common use of chlorine products and historically has controlled widespread waterborne diseases such as cholera and typhoid. Hypochlorites are used as environmental surface disinfectants but residual chlorine may need to be removed to prevent corrosion (28).
Mechanism of Action
Chlorine causes oxidation of proteins and lipids, thereby damaging the cell wall and membrane. Hypochlorous acid is the most active product and even at a neutral pH penetrates the outer cell layers. The moiety disrupts oxidative phosphorylation and other enzymatic activities of the cell. This germicide affects membrane-bound as well as cellular functional proteins, particularly those with sulfhydryl bonds (28), similar to the iodophors. Hypochlorous acid suppressed E. coli growth via complete inhibition of DNA synthesis and viral RNA is degraded into fragments (89). Increased bacterial spore and fungal cell wall permeability has been observed, which causes better biocide penetration and microbe killing (89).
The general mechanisms of chlorine can be summarized as follows:
■ Damage to cell membrane by oxidation of proteins and lipids
■ Inhibition and degradation of DNA and RNA
■ Increased spore and fungal cell wall permeability and subsequent microbe death
Spectrum of Activity
The antimicrobial activity of the halogen releasers and iodine and iodine-containing formulations is similar to chlorine-releasing compounds; these formulations have broad-spectrum activity against gram-positive and gram-negative bacteria, mycobacteria, fungi, and viruses (88). As with most chemical germicides, the activity of iodine and chlorine are dependent on their concentration, the formulation, the temperature, the growth conditions of the microorganism, and the presence of organic and inorganic material.
The sporicidal activity of iodine compounds is significantly less than chlorine-releasing compounds. A concentration of 2,000 to 5,000 ppm potassium iodine is necessary to produce a 5 log10 reduction in B. subtilis spores within 5 minutes. Iodine solutions are somewhat effective in destroying spores of Bacillus anthracis, B. subtilis, Bacillus megaterium, Bacillus mesentericus, Clostridium tetani, and Clostridium welchii (90–92) with prolonged application. Generally, disinfectant formulations of iodophors are weakly sporicidal with prolonged application, whereas antiseptic-containing iodophors are not sporicidal (93).
The activity of iodine against Pseudomonas spp and Legionella pneumophila isolates from a water system was dramatically different if the organisms were grown in water versus rich medium. Water-grown organisms proved more resistant to the iodine (94,95). Studies have also shown that iodine-containing formulations are more effective against vegetative organisms than are chlorine-based formulations (88).
Antimycobactericidal activity of iodine and iodophors are inconsistent (96–98). The following data exemplify the lack of consistency regarding the tuberculocidal activity of iodine and iodophors; hence, these compounds are not considered high-level disinfectants. Some tests showed that very low concentration of free iodine (<0.1%) killed M. tuberculosis with a 5-minute exposure at room temperature (96). In other tests, however, several commercially available iodophor formulations were not effective in killing two strains of M. tuberculosis after a 30-minute exposure (97). A comparison of two different iodine formulations, povidone-iodine (1% titratable iodine) and an iodophor (0.008% titratable iodine), demonstrated that the iodophor was ineffective in killing greater than a 3 log10 CFU M. tuberculosis after 1-minute contact in suspension or dried on a carrier. The povidine-iodine was effective in producing more than a 3 log10 reduction in a suspension test but not when the organism was dried on carriers in the presence of sputum (98). The iodophor product was also ineffective against Mycobacterium smegmatis (99). As a result of these inconsistent tuberculocidal effects, these compounds are not classified as high-level disinfectants.
Chlorine, iodine, and iodine-containing products have good fungicidal and fungistatic properties. They are effective against species of Trichophyton, Monilia, Epidermophyton, Torula, and other genera (88,100).
Iodine and iodophors are effective against a wide range of viruses, including enteroviruses and polio, herpes, vaccinia, rabies, and tobacco mosaic viruses (101–103), with inactivation concentrations of 75 to 150 ppm for polio type 1, coxsackie B1, adenovirus 2, vaccinia, herpes, influenza A, and HIV-1 viruses and 5,000 ppm for feline parvovirus. Complete inactivation of cell-free HIV was accomplished using 0.5% PVP-I with a 30- to 60-second exposure. Quantitative suspension testing has shown virucidal efficacy of povidone-iodine disinfectants against vaccinia, polyomavirus SV40, adenovirus and poliovirus, although more than 60 minutes of exposure was required for the most resistant poliovirus species (104). Some preparations of PVP-I have proven to be active against hepatitis B virus and enteroviruses as measured by the morphologic alteration test, while other preparations had no activity at all (105). In accord with the observation for Pseudomonas spp, this variability may be attributable to the amount of free iodine in the preparation.
Iodine-containing solutions have been primarily used as skin preparation/degerming solutions and surgical hand scrubs. Chlorine antiseptics are not used commonly due to skin irritation. In general, most iodine-containing skin prep and surgical scrubs show rapid reduction in the resident flora of the skin; however, they do not show residual activity. Studies have shown them to be inferior to chlorhexidine gluconate (106). Utilizing the Vienna test method, a 3 log10 reduction was achieved after a 2-minute wash with PVP-I; the same effect was achieved with chlorhexidine gluconate after a 1-minute wash (32). These results were confirmed comparing 14 different iodine preparations (107). In general, PVP-I solutions are more negatively affected by the presence of organic material than are chlorhexidine formulations (108,109).
ALCOHOLS
Alcohols have been used for disinfection of both skin and surface since antiquity (110). In Europe, the use of ethanol as a skin antiseptic has been the standard (110), while in the United States, ethanol is often replaced by antiseptic chemicals such as chlorhexidine, povidone-iodine, and triclosan. Alcohol preparations have been used as hygienic hand disinfectants, surgical hand disinfectants, and skin disinfectants, each having its own set of requirements. In addition, alcohol is often a constituent of combination skin disinfectants. Health care workers should be aware that anionic additives in hand disinfectants containing alcohol may inactivate antibacterial activity and persistence of chlorhexidine (61).
Alcohols have the general chemical formula R–OH, where R can be any organic structure such as an alkyl or benzyl molecule (see Table 12.1). The –OH group is the reactive species and its reactivity is influenced by the attached R group.
Alcohols have been commonly used as hard-surface disinfectants because of their general antimicrobial properties. However, volatility and flammability can be an issue when using alcohols in this manner. The flash points of ethanol and propanols are below 15°C, requiring caution in their use. On the other hand, short-chain alcohols can be suitable as disinfectants because they have unlimited solubility in water, have low toxicity, are fast acting, and are microbicidal. Low-weight alcohols possess less surface tension than water and thus have better wetting characteristics. This is important for antiseptics used on skin or for surface disinfectants used on nonuniform surfaces (110).
Regardless of the application, the order of antibacterial effect of alcohol isomers is n-primary > iso-primary > secondary > tertiary. This review is confined to the use of alcohols as antiseptics (i.e., their use on skin), as that is the primary use in health care today.
Mechanism of Action
It is generally believed that alcohols, like most chemical germicides, act at multiple sites of the cell. The primary mode of action is related to coagulation/denaturation of proteins and solubility of the alcohols in lipids (111,112) (see Table 12.1, Fig. 12.1). In the absence of water, coagulation will not occur. For example, 70% ethanol is more effective than 95% ethanol as a germicide because higher concentrations of alcohol may deprive the bacterial cell of water, thereby inducing an impermeable cell membrane. This activity prevents the penetration of alcohol into the cell (110). Hence, with dry cells, water is needed for antimicrobial activity, though this is not true for moist cells.
Coagulation of cell wall and cytoplasmic proteins occurs after exposure to alcohol disinfectants but does not appear to affect nucleoproteins (113). Coagulation of enzymatic proteins leads to loss of cellular function but there is no specificity with regard to which enzymes are affected. In addition, as the chain length of the alcohol increases, there is further impact on diminishing enzyme function (114). Bacterial enzymes in the cell wall are more easily inactivated than intracellular enzymes (115). Microbe cell lysis following alcohol exposure is likely due to disruption of the cell membrane (110,116,117).
The general mechanisms of alcohols can be summarized as follows:
■ Coagulation/denaturation of proteins and lipids
■ Water-dependent activity due to water requirement to achieve cell membrane permeability
■ Cytoplasmic components coagulate after intracellular concentration
■ Disruption of cell membrane followed by cell lysis
Spectrum of Activity
A review of the in vitro activity against bacterial spores indicates that alcohols are considered to have little killing effect. As a matter of routine procedure, spore preparations are often kept in 95% ethanol to prevent them from contamination by other vegetative organisms. However, other data indicate that some sporicidal activity may be found and is related to the alcohol itself, its concentration, and the species of microorganism (110).
A number of studies document the activity of alcohols against gram-positive and gram-negative organisms (118–120) and against multidrug-resistant pathogens (121–123). Tables 12.9 and 12.10 summarize the antimicrobial activity of a variety of alcohols against common microorganisms using carrier and suspension testing methods.

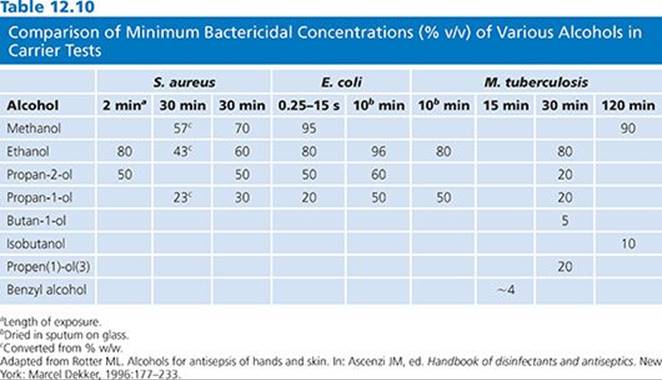
Ethanol is bacteriostatic and inhibits spore growth at concentrations around 10%, is bactericidal at 30% or more, but loses its activity at 90% or more, indicating the need for water for biocidal activity to kill S. aureus as well as other bacterial species (118,124).
Alcohol has in vitro activity against M. tuberculosis and a variety of fungi (110,119,125). Clinical isolates of Mycobacterium have been shown to be resistant to 75% alcohol (the alcohol type was not identified). Mycobacterium chelonae and Mycobacterium nonchromogenicum showed prolonged survival in 75% alcohol (126). Water content is necessary for the effectiveness of this antiseptic (Table 12.11).
The fungicidal activity of short-chain alcohols, mainly ethanol, is well established (110). Most are killed within short exposure periods (5 to 60 minutes) at concentrations ranging from 35% to 96%. Candidaspp and Aspergillus niger appear to be very susceptible, with a minimum effective concentration of 35% at an exposure time of 1 minute.
As with other chemical agents, the virucidal activity of alcohols is generally related to the presence or absence of a lipophilic envelope. Among the enveloped viruses, methanol, at concentrations of 20% to 80%, inactivates viruses in 15 minutes to 24 hours (127,128). Ethanol at concentrations ranging from 50% to 60% inactivates the same viruses in 1 hour (110). The propanols seem more active, since they require lower concentrations to have the same effect with similar exposure times (110,129,130).
The nonenveloped viruses such as picornaviruses, however, are more difficult to kill with longer chain alcohols than with ethanol. Seventy percent ethanol was effective in 2 minutes against poliovirus type 1 (131), though at 25% concentration, a 240-minute exposure was necessary for killing. Ninety-five percent 2-propanol was not effective after exposure for 10 minutes against this virus. Coxsackievirus type B was not killed by 95% 2-propanol after 10 minutes of exposure (130). Echovirus type 6 was inactivated with 50% ethanol in 10 minutes, whereas 90% 2-propanol was not effective. Seventy percent ethanol and 45% 2-propanol were not effective against hepatitis A after 1 minute of exposure (132) but hepatitis B, which is normally considered resistant to many chemical germicides, was relatively sensitive to both alcohols (133,134).
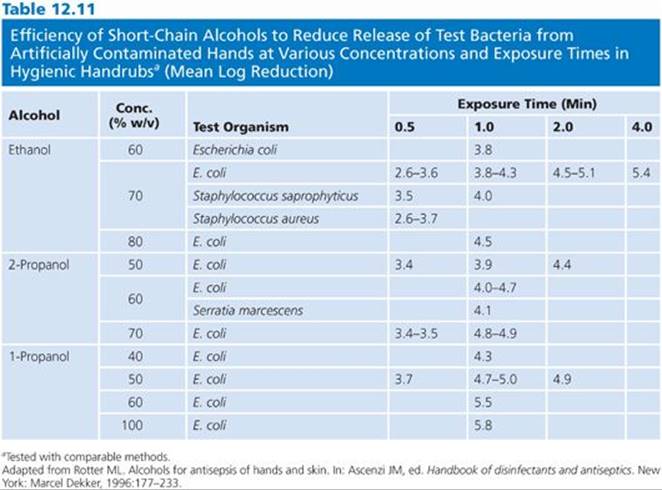
The microflora of the skin can be divided into two groups: the resident flora (more difficult to remove) and the transient flora (easily removed). The resident microflora also replicate on the skin. Health care personnel handwash preparations are designed primarily to remove the transient organisms, whereas surgical scrubs and skin prepping formulations are directed more at reducing the resident flora. It has been demonstrated that although bacteria on a patient’s skin are the main cause of surgical site infections, it is important for health care personnel to further reduce the risk of infection by careful hand hygiene (110). Studies demonstrate that alcohol rubbed on the skin reduce the microbial flora as effectively as a 6-minute scrub with water. Unlike other antiseptics (e.g., chlorhexidine and triclosan), alcohols do not have any residual or substantive properties. Mixtures of alcohols and agents such as chlorhexidine combine the rapid effect of the alcohol with an ingredient with substantive activity (110).
Alcohols have a sublethal effect on skin microflora and allow very slow bacterial regrowth (25,110). Table 12.11 summarizes the data demonstrating the efficiency of short-chain alcohols for reducing bacteria from artificially contaminated hands at various concentrations and exposure times in a hygienic handwash study (110). The data indicate that alcohols are quickly able to reduce bacteria from hands by 2.6 to 5.8 log10, depending on the type of alcohol and the concentration. Concentration and exposure time are critical for both immediate and residual activity. The greatest residual activity is noted when 2-propanol is combined with chlorhexidine.
Health care workers are less likely to transfer gram-negative bacilli to urinary catheters when they use 2-propanol as a handrub rather than washing with soap (135). The data for rhinoviruses is less striking. Seventy percent ethanol impregnated into towels was only slightly more effective than 10% ethanol. Ethanol-2-propanol and 3% benzyl alcohol were only slightly better than 70% ethanol alone (136). Single treatment with an ethanol handwash was ineffective against rhinovirus (137); however, soap and water was effective. Against influenza H1N1, killing was achieved with up to 70% alcohol-based handrubs, with or without chlorhexidine, although soap and water hand hygiene was slightly more effective (138).
ALDEHYDES
Glutaraldehyde and ortho-Phthalaldehyde
Formaldehyde and glutaraldehyde have been used for sterilization and disinfection of medical devices since the 1960s (139). Although formaldehyde is an excellent biocide, its toxicity has limited its use. Glutaraldehyde is used for routine disinfection of medical devices, especially flexible fiber-optic endoscopes and heat-sensitive medical devices. It has excellent biocidal activity and materials compatibility. ortho-Phthalaldehyde (OPA) is a high-level disinfectant. It has the same materials’ compatibility characteristics of glutaraldehyde and enhanced antimicrobial properties. The structure of these agents is shown in Table 12.1. Many other aldehydes have antimicrobial activity but have not been developed into commercial products and hence will not be discussed here.
Mechanism of Action
Several aldehydes have antimicrobial properties, but the most widely used are glutaraldehyde and formaldehyde. Benzaldehyde, succinaldehyde, malonaldehyde, glyoxal, and several of the β-unsaturated aldehydes (e.g., cinnamaldehyde) also have antimicrobial activity.
The antimicrobial activity of aldehydes in general is based on the reactivity of the aldehyde group (140) and its ability to undergo alkylation reactions. Reactivity of this group can be changed by other functional groups on the molecule. Aldehydes tend to become hydrated in aqueous solutions and in equilibrium with the free aldehyde form, which is believed to be the active moiety. In the case of glutaraldehyde, the monomeric-free aldehyde molecule is in equilibrium with the cyclic hemiacetal and acetal polymers, together coexisting with smaller quantities of mono- and dihydrates. This equilibrium appears to be temperature sensitive, since at higher temperatures, there is a shift to the more active monomeric form. The effect of pH on aldehydes is well described. The reaction of glutaraldehyde with protein increases as the pH rises from 4 to 9 (141). Glutaraldehyde activity is greater at a basic pH of 9 to 10, possibly because condensation reactions are catalyzed better at an alkaline pH (141).
Amines and sulfhydryl groups react most strongly with aldehydes. Cellular constituents involved most often are both enzymes and structural proteins and nucleic acids. Treatment of Micrococcus lysodeikticuscells with glutaraldehyde prevented the release of selective enzymes from the periplasmic space, indicating interaction of glutaraldehyde with the outer cell surface (142). In poliovirus type 1 and echovirus type 25, glutaraldehyde reacts with lysine residues in the protein capsid (143). Enzymatic function via DNA polymerase was affected when the virus was exposed to a glutaraldehyde disinfectant (144). Formaldehyde is known to react with amino groups to bring about intermolecular cross-linking (145) and although it reacts with both RNA and DNA, its reaction with RNA is stronger (146). Glutaraldehyde, like formaldehyde, can cause intramolecular and intermolecular cross-linking of molecules. Antimicrobial properties of OPA involve binding to membrane receptors followed by entrance of the biocide into the cell through the permeable membrane. OPA then compromises cell cycle function through disruption of DNA and RNA (147). Transport of low-molecular-weight amino acids in glutaraldehyde-treated E. coli was reduced about 50%. One would expect much less transport if this was the only mechanism of action (148). In a comparative study of mechanism of action of glutaraldehyde and of OPA on mycobacterial cells, the data suggest that the cross-linking effect of glutaraldehyde on the cell membrane is the mechanism by which glutaraldehyde exerts its antimicrobial action and that the more lipophilic OPA molecule is more efficient at crossing the lipid-rich cell surface of the mycobacterial cell and exerting its action internally (149).
Investigations into the sporicidal activity of glutaraldehyde indicate that the spore coat is protective and that the glutaraldehyde must be alkalinized in order to penetrate this protective barrier (150–152).
The general mechanisms of aldehydes can be summarized as follows:
■ Interaction with cell membrane and cytoplasmic proteins, especially those with sulfhydryl moieties
■ Penetration into cells is pH dependent, with aldehydes working better at basic pH
■ Intramolecular and intermolecular cross-linking of molecules
■ Compromise of cellular function by disrupting DNA and RNA
Spectrum of Activity
Glutaraldehyde was first proposed as an antimicrobial in 1962 (139) as an alternative to formaldehyde sterilization of sutures. The sporicidal properties of alkaline glutaraldehyde were demonstrated in 1963 (153), making this form preferable as a chemical sterilant. OPA is a commercially available high-level disinfectant. Since the only aldehydes commercially available for hospital use in the United States are glutaraldehyde and OPA, the spectrum of activity of other aldehydes, although of interest, is not dealt with here.
The literature describes the effectiveness of glutaraldehyde against gram-positive and gram-negative bacteria. Rubo et al. (154) demonstrated the rapid bactericidal activity of 0.02% glutaraldehyde against Staphylococcus aureus, E. coli, and Pseudomonas aeruginosa. In a 20-minute exposure to this low concentration, inactivation of 104 cells or more was observed. Pseudomonas aeruginosa appeared to be the most resistant of the three organisms tested. Additional work by Borick (155) demonstrated rapid kill (less than 1 minute) of E. coli, Pseudomonas aeruginosa, Serratia marcescens, Proteus vulgaris, and K. pneumonia by 2% glutaraldehyde. Helicobacter pylori was rapidly killed by 0.5% glutaraldehyde with exposure as short as 15 seconds (156). OPA has been shown to be effective against both vancomycin-resistant enterococci (VRE) and MRSA (157).
Several strains of atypical mycobacteria (158–162) were assessed for susceptibility to glutaraldehyde. In evaluating a modified in-use method, it was shown that exposure of strains of Mycobacterium avium-intracellulare, Mycobacterium gordonae, Mycobacterium fortuitum, and M. chelonae to 1% glutaraldehyde for 15 minutes resulted in a 4 log10 reduction, and exposure to a 2% solution for 1 minute resulted in 100% kill (160). In another study, 2% glutaraldehyde tested against several of the same species in a quantitative suspension and carrier test was shown to reduce their populations 5 log10 within 30 minutes in the presence of organic soil and hard water (161). Further work by the same group showed that endoscopes artificially contaminated with M. tuberculosis and M. avium-intracellulare in sputum and endoscopes precleaned using a neutral soap could be disinfected with 2% glutaraldehyde in 10 and 20 minutes, respectively (162). The data indicate that there is a range of times for the inactivation of the various species. Collins (160) found that the most resistant species to glutaraldehyde were M. gordonae and members of the M. avium-intracellulare group (Table 12.12).
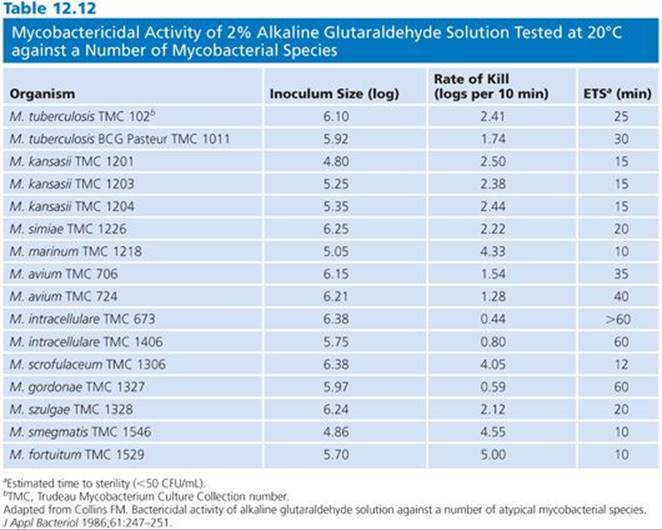
Microorganisms isolated from water systems or grown in water are much more resistant to the action of disinfectants than bacteria grown in laboratory media (163,164). Studies have evaluated a variety of disinfectants against several strains of atypical mycobacteria, including glutaraldehyde-resistant strains isolated from washer disinfectors. As expected, the washer disinfector isolates were extremely resistant to glutaraldehyde but notably, both acidic OPA and alkaline OPA, under dirty and clean conditions, were effective (158). The mycobacteria are a heterogeneous group of organisms but the one thing they have in common is a cell wall with high lipid content. This makes them less susceptible to hydrophilic molecules, hence less vulnerable to many disinfectants. The surrogate organism used to determine tuberculocidal activity is Mycobacterium bovis var BCG. Collins and Montalbine (159) examined the activity of glutaraldehyde against a variety of mycobacteria, including the virulent strain M. tuberculosisH37Rv.
OPA has several advantages over glutaraldehyde, including its superior activity against mycobacteria. Based on the claims for high-level disinfection, a 0.55% solution of OPA is effective within 5 minutes, whereas a 2.5% glutaraldehyde solution requires 45 minutes to achieve the same 6 log10 reduction in M. bovis (158).
In general, aldehydes are less effective against bacterial spores than vegetative forms of the same organism and other vegetative organisms. Glutaraldehyde is the only aldehyde with substantial activity against spores. A 2% alkaline glutaraldehyde solution has the same activity as an 8% formaldehyde solution (153,165) and 10 times the activity of a small aldehyde, glyoxal (ethanedial) (139). The sporicidal activity is directly related to the pH of the solution; alkaline solutions have been shown to be more sporicidal. When bicarbonate is added to glutaraldehyde, substantial increases in the sporicidal activity result (139). It appears that bicarbonate affects the spore and not the glutaraldehyde molecule, as pretreatment of spores with bicarbonate renders them sensitive to acidic glutaraldehyde (166).
C. difficile is a clinically important spore-forming bacterium, especially in hospitalized patients and following the use of certain antibiotic therapies (167–170). Data indicate an in vitro susceptibility of C. difficile spores to glutaraldehyde with short exposure periods of 5 to 30 minutes for 0.1% to 2.0% glutaraldehyde. Longer exposure times are required to kill B. subtilis and Bacillus stearothermophilus spores (171).
A study evaluating the performance of 2% glutaraldehyde against C. difficile in situ corroborates the in vitro data (172). Endoscopes contaminated with C. difficile were effectively disinfected in 5 minutes. A comparison of glutaraldehyde and of OPA indicates that glutaraldehyde is considerably more sporicidal. Two percent and 3% glutaraldehyde solutions achieved a 6 log10 reduction in spores of B. subtilis in 6 and 3 hours, respectively, while 0.3% and 0.55% OPA required 72 and 48 hours, respectively. With spores of C. tetani, 2% and 3% glutaraldehyde achieved a 2 log10 reduction in 2 minutes, and 0.3% and 0.55% OPA achieved the same results in 20 minutes and 5 minutes, respectively. For spores of C. difficile, 2% and 3% glutaraldehyde achieved a 3 log10 reduction in 15 seconds, whereas 0.3% and 0.55% OPA achieved the same level of sporicidal activity at 2 minutes and 1 minute, respectively (173).
Fungi, as yeasts and molds, are generally susceptible to glutaraldehyde at concentrations marketed as high-level disinfectants. The effect of pH again has been demonstrated. A 0.5% solution of alkaline glutaraldehyde was shown to be more effective than an acidic solution at the same concentration at inhibiting spore production and growth of A. niger (174). A 2% acidic glutaraldehyde solution, however, was found to be effective in 15 minutes at 25°C against a wide variety of fungi except A. niger, which required 30 minutes for complete kill. The MIC for acidic glutaraldehyde was five times higher than for alkaline glutaraldehyde in inhibiting the growth of S. cerevisiae (175). Other work has shown glutaraldehyde to be potent against a wide variety of fungi, including Trichophyton interdigitale, Microsporum gypseum, C. albicans, Mucor hiemalis, Rhizopus stolonifer, Penicillium chrysogenum, and Byssochlamys fulva (176). A 10% alkaline glutaraldehyde solution was shown in vitro to eradicate Trichophyton mentagrophytes and Cephalosporium and Fusarium spp (177). In vivo activity of glutaraldehyde was demonstrated in the treatment of onychomycoses (178). Isenberg et al. (179) demonstrated the effectiveness of 2% alkaline glutaraldehyde against Candida albicans in normal hospital practice. The data in Table 12.13 indicate that, with normal hospital use of a commercially available glutaraldehyde, the activity of the solution was maintained for 2 to 4 weeks following in-use stress.
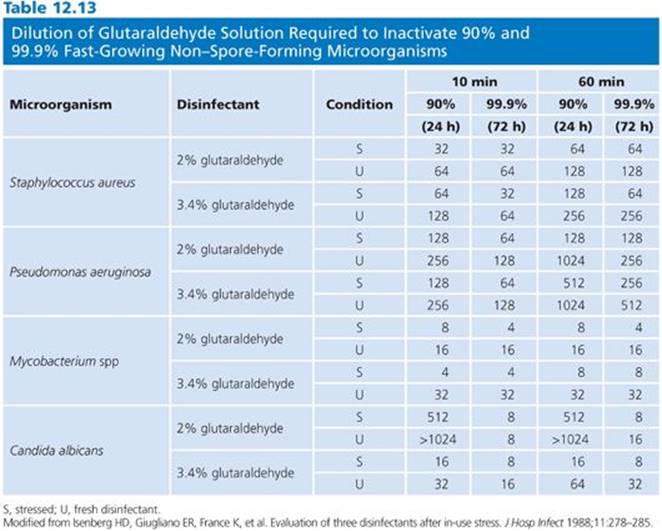
Glutaraldehyde has proven virucidal activity. Glutaraldehyde is active against enveloped and nonenveloped viruses, the former being more susceptible. Poliovirus type 1, coxsackievirus type B1, and echovirus type 6 were more difficult to inactivate, requiring greater than 1% glutaraldehyde, than were herpes simplex virus, vaccinia virus, Asian influenza virus, and adenovirus. (Although adenovirus is classified as a nonenveloped virus, it does contain lipids absorbed to its protein surface and they are considered an integral part of the virus structure, as with enveloped viruses.) The enveloped viruses and adenovirus required 0.2% or more glutaraldehyde with a 1-minute exposure at room temperature for inactivation (143). Isolates of echovirus showed differences in susceptibility to glutaraldehyde. Table 12.14 lists the viruses susceptible to glutaraldehyde (180–196).
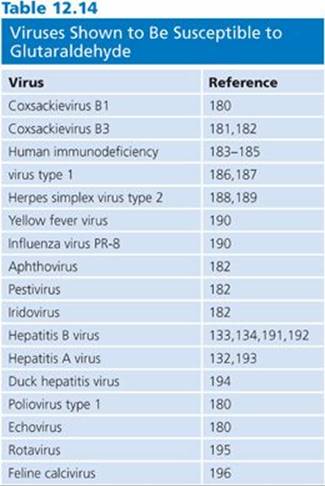
After an exposure for 5 minutes at 20°C, OPA is effective against bovine viral diarrhea virus, a surrogate test organism for hepatitis C virus (197), and against duck hepatitis B virus, a surrogate test virus for human hepatitis B virus (198). Organic material, which affects the activity of many chemical germicides, has little effect on the virucidal activity of glutaraldehyde (181,182,189,190,195). Saitanu and Lund (181) present data that indicate improved activity of glutaraldehyde in the presence of organic material (10% horse serum and 20% mouse droppings). Glutaraldehyde was also shown to retain its activity against cell-associated HIV in the presence of blood (187).
PEROXYGEN COMPOUNDS: HYDROGEN PEROXIDE AND PERACETIC ACID
The two peracids that are the most commonly used for the purposes of disinfection and/or sterilization are hydrogen peroxide and peracetic acid. Both of these compounds belong to the group known as peracids or peroxygen compounds.
Hydrogen peroxide has been used for its antiseptic properties since the 1800s. It is most useful for applications where breakdown into nontoxic by-products is crucial (199). Originally, hydrogen peroxide was used for the preservation of milk and water and for the sterilization of certain beverages. As an antiseptic, it has been used in the treatment of periodontal disease and endodontic therapy as well as for oral topical applications. As a disinfectant, it is used at various concentrations to disinfect materials such as drinking water, medical equipment, and septic tank waste. It was the first nonthermal disinfectant cleared by the FDA for use on contact lenses (200). The general structure of these agents is shown in Table 12.1.
Mechanism of Action
Hydrogen peroxide is produced in phagocytes, where it kills bacteria. The activity of commercially produced hydrogen peroxide is less affected by pH than that of many other disinfectants such as phenols, organic acids, and glutaraldehyde (201).
Peracetic acid may be a more potent antimicrobial than hydrogen peroxide. Peracetic acid has the added advantage of being more lipid soluble and not susceptible to decomposition by catalase and peroxidase (202), which effectively neutralize the action of hydrogen peroxide. Like hydrogen peroxide, it has been used for the disinfection and sterilization of medical devices (e.g., hemodialyzers) and environmental surfaces. It is more resistant than hydrogen peroxide to the neutralizing effect of organic material (25).
The hydroxyl radical is an extremely potent oxidant and is responsible for the killing action of peroxide (203). The free hydroxyl radical reacts with essential components of the cell such as lipids, proteins, and nucleic acids. Antimicrobial properties of hydrogen peroxide may be due to the oxidation of double bonds in proteins, lipids, and surface membranes. However, it has other known effects, such as dissociation of 70S ribosomal subunits (see Fig. 12.1). These effects cause cell surface changes and cleavage of the DNA backbone and hence damage to spore DNA (204). Investigations suggest that the oxidizing radical is the ferryl radical formed from DNA-associated iron. Research also suggests that selective oxidation of certain proteins occurs and that vapor-phase hydrogen peroxide is a more potent oxidizer of protein than liquid-phase hydrogen peroxide (199).
The mechanism of action of peracetic acid is not as well studied but it functions as an oxidizing agent. Therefore, its mechanisms include denaturing proteins, disrupting cell membrane permeability, and oxidizing sulfhydral bonds in proteins and enzymes (4,25,205). Decomposition of peracetic acid produces the highly reactive molecule singlet oxygen and it has been proposed that this moiety is responsible for peracetic acid’s activity (206). Malachesky (207) reviewed the chemical reactions of peracetic acid. Many of the groups that peracetic acid reacts with are present throughout living cells; therefore, the mechanism of action is probably not specific to any one cell structure.
The general mechanisms of peroxides can be summarized as follows:
■ Free hydroxyl radical reacts with essential components of the cell such as lipids, proteins, and nucleic acids
■ Dissociation of 70S ribosomal subunits
■ Cell surface changes and cleavage of the DNA backbone
■ Oxidation causing denaturation of proteins, disruption of cell membrane permeability, and sulfhydral bonds in proteins and enzymes
Spectrum of Activity
A number of investigators have looked at the activity and disinfecting properties of 3% hydrogen peroxide expressed as D-values. Table 12.15 lists these values. Bacteria show rapid disinfection; however, the two fungal species tested show more resistance to 3% peroxide. The data reveal discrepancies between the two studies with regard to the resistance. C. albicans was very resistant in both studies, indicating that longer disinfection times would be required for this organism. D-values for organisms on the surfaces of contact lenses were lower (201). Tables 12.16 and 12.17 indicate the lethality of peroxide at concentrations less than 3% toward a number of bacteria, fungi, viruses, and spores.



Peroxide has a wide spectrum of activity. Its activity is generally better against gram-negative than gram-positive organisms. In addition, viruses are extremely susceptible, being inactivated at concentrations of 30 ppm or less, although some viruses required extended periods of contact. Higher concentrations or temperature would presumably speed the action. Most bacteria are easily killed with relatively low concentrations of peroxide and in a relatively short period of time. Anaerobes are even more susceptible because they lack the enzymes catalase, peroxidase, and superoxide dismutase. However, to kill spore-forming bacteria such as B. subtilis and C. sporogenes, 6% hydrogen peroxide or 2% glutaraldehyde solutions applied for 6 and 10 hours, respectively, are needed for effective sporicidal activity. However, at lower dilutions, even such prolonged hours of exposure are not efficacious (208). Alternatively, a dilution of 0.2% peracetic acid at 50°C in the presence of 5% serum and hard water shows rapid activity against vegetative bacteria, fungi, and spores (207).
Peracetic acid is effective against M. tuberculosis, M. smegmatis (207), and a variety of clinical isolates of Mycobacterium and Cryptosporidium spp (209). Table 12.18 presents a comparison between peracetic acid and two other disinfectants commonly used on environmental surfaces, chlorine and benzalkonium chloride. The two other disinfectants require a much longer duration of exposure to achieve the same result as peracetic acid. Combinations with other biocides may also have synergistic effects against various microorganisms (28).

New trends in peroxide disinfectants include low-temperature sterilization technology, hydrogen peroxide gas plasma, vaporized hydrogen peroxide, and ozone. Ozone as a disinfectant has increasing applicability in hospital settings. Newer generators permit maintenance of adequate ozone concentrations and temperature control which improve its functionality in hospital disinfection. Ozone often plays a role in improving the efficacy of mixed products, which contain chlorine, for example, thereby exerting increased antimicrobial action (28).
Both hydrogen peroxide and peracetic acid have been commercialized in liquid and vapor forms. It is generally accepted that the vapor phase of each compound possesses great antimicrobial activity. Low-temperature sterilizing systems using vaporized hydrogen peroxide has been shown to be very effective (210). The difference in effectiveness may be due to oxidative structural changes in the gaseous as compared to the liquid forms of hydrogen peroxide (205). In the liquid phase, each compound is unstable and requires careful handling and storage to ensure it maintains its level of antimicrobial activity. Prions may be sterilized more readily by vaporized hydrogen peroxide generators (211). These generators produce gas, which has greater antimicrobial activity at lower concentration than the liquid form of hydrogen peroxide. Vaporized products can be used for cleaning critical environments and rooms.
PHENOLS: PHENOLS, TRICLOSAN, AND HEXACHLOROPHENE
Phenol use as antiseptics and disinfectants dates to the application of coal tar poultices. However, it was the use of carbolic acid (currently referred to as phenol ) by Lister as an antiseptic during surgery that demonstrated this compound’s antimicrobial potential. The phenolic group comprise compounds whose basic structure is phenol. Phenol itself is the simplest of the group. The more complex compounds include p-tert-amylphenol and phenylphenols such as hexachlorophene and triclosan (2,4,4′-trichloro-2′-hydroxydiphenyl ether) (see Table 12.1). In general, phenolic compounds are used in the hospital setting as low- to intermediate-level disinfectants. Effectiveness of other disinfectant is compared to phenol as an industry standard (phenol coefficient method; see Table 12.4). They are primarily used on environmental surfaces. Other phenolics have been used in skin antiseptic formulations. Triclosan is commonly used in home products (toys, kitchen utensils, etc.), cosmetics, oral hygiene, and dermatologic preparations as well as health care personnel handwashes. Phenolic disinfectants should not be used to disinfect critical instruments because they are not sporicides and exhibit inadequate tuberculocidal activity. Treatment of semicritical devices is also limited because of the difficulty rinsing them from many materials as well as patient tissue irritation caused by residuals (82).
Mechanism of Action
The mechanism by which phenolic compounds exert their antimicrobial actions is not fully elucidated. The free hydroxyl group is the reactive site of the molecule. Alkyl or halogen substitution affects the reactivity of the hydroxyl group. The hydrophobic nature of the polar group contributes to the membrane-active properties of phenolic compounds.
In general, phenolics exert their biocidal activity by binding to the cell surface, targeting the cell membrane. Subsequent to binding, interference with membrane-associated functions takes place (212). It is hypothesized that binding to the membrane results in inhibition of respiratory activity, substrate oxidation, and transport processes (213). Hexachlorophene inhibits the membrane-bound part of electron transport near the terminal acceptor (214). Uncoupling of oxidative phosphorylation is supported by studies on the effects of 4-ethylphenol on E. coli and of fentichlor on E. coli and S. aureus (212,215–218).
Phenolic compounds affect cell membrane permeability. A strong correlation exists between the loss of K+ and cell viability in Serratia marcescens exposed to phenol (219). Treatment of Streptococcus faecalis with 1% phenol produced leakage of amino acids consistent with cell lysis (220). Leakage of 14C-labeled glutamate, 14C-adenosine, and 32P-phosphate from cells exposed to several phenolic compounds further substantiates this observation (221–224). The extent of release directly correlates with the concentration of the biocide. Early events were shown to be reversible, but ultimately, leakage led to cell death. The high lipid content of the mycobacterial cell wall is thought to be this organism’s target for phenols. In mycobacteria, phenols are not particularly sporicidal but have been shown to be sporostatic, inhibiting germination and outgrowth (225). Phenol appears to bind loosely to sites on the spore surface, exerting its inhibitory but reversible effects (226). Inhibition of cellular systems by phenolic compounds has also been reported, suggesting that after penetrating the cell membrane and entering the cytoplasm, a secondary site of action may be cytoplasmic enzymes. This is further suggested by the observation of coagulation of cytoplasmic proteins in phenol-treated cells (227,228). Treatment of Staphylococcus aureus with a 1:1,000 dilution of phenol also inhibited activation of several enzymatic pathways, although to varying degrees (229,230). Succinate dehydrogenase, a cytoplasmic enzyme, was inactivated by 4-butyl phenol and hexylresorcinol at concentrations higher than those required for microbicidal activity (114).
The phenol triclosan also interferes with the function of the cellular membrane. It inhibits uptake of certain amino acids (231). Activity of triclosan is also related to its effect on fatty acid synthesis (FAS). Bacteria utilize dissociative FAS or type II FAS (FAS-II), as opposed to the single multifunctional enzyme system in associative FAS or type I FAS (FAS-I) present in animal cells and fungi. McMurray et al. (232,233) showed that a mutation in the gene responsible for NADH-dependent trans-2-enoyl-acyl carrier protein reductase of bacterial FAS resulted in an increase in the tolerance of M. smegmatis and E. colito triclosan. The fabI gene encodes this enzyme, which catalyzes the last step in the elongation of fatty acid biosynthesis. Triclosan is known to be very effective against S. aureus, including MRSA, by also inhibiting de novo fatty acid biosynthesis (234). The fabI gene product is also the biocide target in this organism (235,236). Additionally, the FAS-II system has been identified in Plasmodium falciparum(237). Triclosan analogs were shown to bind to the protein encoded for by the enoyl acyl carrier protein reductase gene from P. falciparum. Inhibition is thought to occur by mimicking the natural substrate of the reductase enzyme. The three-dimensional structure of this enzyme binds to triclosan, which further substantiates this observation (238). These studies suggest mechanisms other than cell membrane damage are involved with triclosan activity. At higher concentrations, as available in commercial products, cell membrane disruption may be the primary mechanism of action. Very little information appears in the literature concerning the specific mechanism of action of phenols against fungi and viruses.
An analysis and review of the structure–activity relationship of phenolic compounds reveals the general mechanisms of phenols, which can be summarized (220) as follows:
■ Halogen substitution intensifies the microbicidal activity.
■ Introduction of aliphatic or aromatic groups into the nucleus further increases bactericidal activity.
■ Aliphatic chains intensify the bactericidal activity to a greater degree than branched chains or 2-alkyl groups of the same total number of carbon atoms.
■ 2-Alkyl derivatives of 4-chlorophenol are more germicidal that 4-alkyl derivatives of 2-chlorophenol.
Spectrum of Activity
Most commercially available phenolic products are a mixture of two or more phenolic chemicals. Substitution of the ring structure of phenol changes the reactivity and activity of that base compound. Table 12.19 shows the activity of commonly used phenolic compounds. Based on this table, it appears that Pseudomonas is more resistant than other organisms, whereas the other gram-negative and gram-positive bacteria and fungi are inhibited at relatively low concentrations. Studies with parachlorometaxylenol (PCMX) used in topical preparations confirm this (239). However, higher resistance, as demonstrated by MIC and MBC data, may not be important, as most commercially available products are formulated at much higher concentrations.

Improved antimicrobial activity has been achieved with the combination of ortho-phenylphenol and ortho-benzyl-para-chlorophenol, which is commonly used in hospital disinfectants. There is still little evidence to suggest that these compounds have substantial activity against mycobacteria, including M. tuberculosis and M. smegmatis (98,99,240). For this reason, they are generally not classified as high-level disinfectants. Commercially available phenolic disinfectants completely inactivated cell-free HIV and cell-associated HIV in culture medium, but when suspended in blood, this virus was not completely inactivated when exposed to the same formulations (241,242). Phenolic compounds generally require long exposure times and high concentrations to be effective against hepatitis B virus (105).
Triclosan has been shown to have activity against a wide variety of microorganisms, including anaerobic bacteria including multidrug-resistant S. aureus, and fungi (243). Determination of MIC with clinical strains of S. aureusindicated that these strains were highly susceptible (MIC90 = 0.12 μg/mL) compared with S. epidermidis (MIC90 = 8 μg/mL) (244). Triclosan was recommended for the control of MRSA outbreaks after publication of several reports of its successful use in a neonatal nursery and cardiothoracic surgical unit (245). The daily use of triclosan in patient baths resulted in MRSA with significantly higher MIC to the germicide. The transfer of resistance was associated with plasmid-mediated mupirocin resistance (246). When 1% triclosan was introduced into hospital units, there was a reduction in MRSA infections over a 7-week period, from 3.4 to 0.14 cases per week, but there was no change in the control (triclosan-free) hospital units (247).
Like chlorhexidine, triclosan binds to the skin and exhibits moderate substantivity, hence its use in surgical preparation formulations and health care personnel handwash preparations (248,249). Triclosan, along with PCMX, was shown to be less effective as a skin antiseptic than formulations containing chlorhexidine.
Studies on oral preparations containing triclosan indicate that in an in vitro system simulating a periodontal disease–like state, exposure of organisms to triclosan or triclosan monophosphate (a more soluble form) caused gram-negative anaerobic organisms to be inhibited to a greater extent than gram-positive organisms (250,251). In studies utilizing pure cultures of oral microorganisms, the results indicate the same trend, with streptococci and lactobacilli being the least susceptible and the organisms Neisseria subflava, Prevotella nigrescens, and Porphyromonas gingivalis being highly susceptible to 0.6 mg/mL triclosan (252).
QUATERNARY AMMONIUM COMPOUNDS
Quaternary ammonium compounds (QACs) are widely used as disinfectants. However, by the 1990s, the CDC advised elimination of QACs as antiseptics because of several outbreaks of infection associated with in-use contamination (253,254). As with several other products (e.g., phenolics and iodophors), disinfectant-resistant gram-negative bacteria have been found to survive or grow in QAC (255). Several reports have documented the association of nosocomial infections associated with contaminated QAC used to disinfect patient care supplies or equipment such as cystoscopes or cardiac catheters (256,257). The quaternaries are good cleaning agents, but use with hard water can influence their antimicrobial effectiveness. Materials used to apply the QAC, such as cotton and gauze pads, can make them less microbicidal, as these materials absorb the active ingredients.
Chemically, the quaternaries are organically substituted ammonium compounds in which the nitrogen atom has a valence of five (see Table 12.1). Four of the substituted radicals (R1–R4) are alkyl or heterocyclic radicals of a given size or chain length, and the fifth (X−) is a halide, sulfate, or similar radical. Each compound exhibits its own antimicrobial characteristics. The chemical names of QACs used in hospitals include alkyl dimethyl benzyl ammonium chloride, alkyl didecyl dimethyl ammonium chloride, and dialkyl dimethyl ammonium chloride. Fourth-generation QACs, referred to as twin-chain or dialkyl quaternaries (e.g., didecyl dimethyl ammonium bromide and dioctyl dimethyl ammonium bromide), purportedly remain active in hard water and are tolerant of anionic residues (258).
Mechanism of Action
The bactericidal action of the quaternaries has been attributed to the inactivation of energy-producing enzymes, denaturation of essential cell proteins, and disruption of the cell membrane. In bacteria, QACs may cause structural and functional changes in the cell wall, release of wall components, cell lysis, inhibition of membrane ATPase, and interactions with negatively charged polar groups of phospholipids (259). The general mechanism of action of QAC compounds are summarized as follows:
■ Penetration through the cell wall
■ Binding to cell membrane lipids and proteins
■ Disruption of cell membranes and, with higher concentrations, cellular leakage
■ Intracellular damage to proteins and DNA
■ Lysis of cell wall components by autoenzymes
■ Loss of structural organization of the microbial cell (112,258,259)
Spectrum of Activity
The quaternaries are commonly used in ordinary environmental sanitation of noncritical surfaces such as floors, furniture, and walls. Manufacturers’ data sheets and results from published studies indicate that the quaternaries sold as hospital disinfectants are generally fungicidal, bactericidal, and virucidal against lipophilic viruses; they are not sporicidal and generally not tuberculocidal or virucidal against hydrophilic viruses. Gram-positive bacteria are generally more susceptible than gram-negative bacteria, thought to be due to the outer membrane of gram-negative bacteria, absent in gram-positive organisms. QACs are effective against S. aureus and protozoa, plasmodia in particular (259). Best et al. (98) demonstrated the low level of effectiveness of QACs against mycobacteria.
Attempts to reproduce the bactericidal and tuberculocidal results claimed by manufacturers using the AOAC tests with a limited number of QACs have not demonstrated consistent microbe killing (260,261). QACs exhibit significant antimicrobial activities but to varying degrees based on the hydrophobic group at the 4-amino side chain position. QACs having larger hydrophobic groups are significantly more effective against E. coli and S. aureus(262).
RESISTANCE
Mechanisms of resistance to antibiotics are well understood. The mechanisms generally fall into one of three categories: (a) altered targets, (b) production of enzyme(s) that degrade the active molecule, and/or (c) decreased ability to take up the compound. Resistance to many antibiotics is linked to the cell’s chromosomes or a plasmid (263,264). However, the actions of chemical antiseptics or disinfectants are, in most instances, not specific for a target molecule. In fact, these biocides may act at several sites in the cell (see Fig. 12.1; Table 12.20). The response of cells to disinfectants is not likely to be as specific as the response to antibiotics. Organisms may develop resistance to various chemicals following heavy use and then revert to normal responsiveness after removal of the active ingredient. There is some evidence that there is an association between plasmid-linked antibiotic resistance and resistance to disinfectants (265–267) (Table 12.21). While some researchers are not convinced that the use of germicides has significantly increased the resistance of microorganisms to antibiotic therapies (268,269), others have pointed to disinfectants and antiseptics as a cause of antibiotic resistance (270). Increased resistance to germicides used in the sanitization of environmental surfaces has, at times, been implicated, as in the outbreak of antibiotic-resistant Listeria monocytogenes in food processing plants (271).


McDonnell and Russell (25) emphasized that resistance to disinfectants and germicides does not necessarily result in inadequate germicidal action. Unlike antibiotics, where an increase in an MIC can have a significant negative therapeutic impact, increased MIC values for antiseptics and disinfectants may not have a noticeable effect. This is because antibiotics work on specific targets within the cell yet antiseptics and disinfectants generally work on the principles of chemical reactions (e.g., oxidation, alkylation, and detergency), hence many cell components may be susceptible to the action of these germicides.
Resistance of microorganisms to chemical germicides can be the result of intrinsic or acquired resistance. Acquired resistance develops through a change in the genetic makeup of the cell, either by mutation or by acquisition of plasmids. Intrinsic resistance is demonstrated in naturally occurring isolates. This is genetically controlled and may be constitutive (i.e., the normal state of the cell) or inducible or adaptive (i.e., in response to an agent).
Intrinsic Resistance
Intrinsic resistance may represent adaptive changes in the cell wall and/or outer membrane structure in response to exposure to germicides. Intrinsic resistance to chemical germicides may occur as a result of several different mechanisms (see Table 12.20). One prominent mechanism is reduced permeability to the agent due to changes in the exterior of the cell or spore (cell wall and/or outer membrane) (272). Intrinsic resistance in this scenario may be due to the natural state of the organism, as with the impenetrable high lipid content in the cell wall of some organisms, making them resistant to antibiotics and germicides (273). Efflux mechanisms (274–277) that prohibit or reduce internal concentrating capability of the germicide are another potential and relatively common pathway of resistance. A third mechanism, degradation of the germicide by enzymatic activity and rendering of the active ingredient ineffective, has occasionally been reported as a mechanism (278). Finally, increased microbe resiliency due to phenotype switching when living in certain growth conditions, an intrinsic virulence factor for some organisms, may also change susceptibility to some disinfectants such as for Candida species to chlorhexidine (100).
Biofilm production has been associated with intrinsic resistance of organisms to germicides. Biofilm production is usually associated with adherence of microorganisms to a surface and production of extensive layers of polysaccharide exopolymer (279,280). Biofilms have been found on devices and are a major component of dental plaque. However, there is no evidence that cells form biofilms in response to germicides. The resistance due to biofilms is primarily a physical phenomenon in which cells are protected from chemical germicides by the extracellular material, which either inhibits access of the biocide to the cells or interact with the biocide to effectively neutralize it. Although the biofilm formed by microorganisms may be resistant to a given germicide, once the biofilm is disrupted and the cells are subcultured, they do not show the same level of resistance. This indicates the mechanism of resistance of biofilm to be primarily physical (281). There have been reports of glutaraldehyde-resistant strains of M. chelonae isolated from automated endoscope reprocessors and processed scopes (282–284). This was due to the formation of biofilms on the surfaces of the tubing in the machines. The strain, which had never been exposed to disinfectants, was very sensitive to all the disinfectants tested, with the exception of the peroxygen. The isolates from the scope disinfector, however, were less susceptible to the peroxygen- and chlorine-releasing compounds and were very resistant to glutaraldehyde, with some strains showing a less than 1 log10 reduction with a 60-minute exposure to 2% glutaraldehyde (285). It was suggested that glutaraldehyde may have selected for the growth of resistant strains of M. chelonae (284).
Bacterial spores, known to be resistant to many chemical and physical agents, exemplify intrinsic resistance by impermeability. The spore coat has also been shown to be the structure responsible for resistance to OPA, for example (286). Bacillus and Clostridium spores are known to be the most resistant to antiseptics and disinfectants (25) and, as stated earlier, resistant C. difficile is a major cause of infections in hospitals and other health care facilities. Some chemical germicides are known to be sporicidal (glutaraldehyde, peracetic acid, and peroxide), but they require high concentrations and/or long periods of contact and/or high temperatures to be effective (Table 12.22) (287). Russell and collaborators have elucidated the development of resistance to a number of germicides during sporulation (57,288,289). As sporulation proceeds (developmental stages 0 through VI), spores become increasingly resistant to germicides in the following order: formaldehyde, sodium lauryl sulfate, phenol, m-cresol, chlorocresol, chlorhexidine gluconate, cetylpyridinium chloride, sodium dichloroisocyanurate, and glutaraldehyde. Resistance to germicides develops primarily during stages IV through VII, correlating with the start of cortex development, coat synthesis, maturation, and release of mature spores. Spore coat formation plays a significant role in the resistance to chlorine-containing compounds (290–292) and formaldehyde. Cortex formation is critical in conferring resistance to QACs, chlorhexidine, and glutaraldehyde (289,293). Other factors involved in the resistance of spores to chemical agents include conformational changes in the inner cell membrane conferred by small multi-drug resistance proteins (294).
Mycobacteria as a group are more resistant than other vegetative bacteria and less resistant than bacterial spores. Mycobacterium spp represent a unique class of bacteria having high lipid content in their cell walls. It has been proposed that these lipids interfere with the uptake or penetration of hydrophilic molecules into the cell. Support for this hypothesis can be found in the relative sensitivity of mycobacteria to disinfectants with varying hydrophilic properties. Two aldehydes, glutaraldehyde and OPA, have vastly different effectiveness against mycobacteria. Glutaraldehyde is a highly water-soluble compound, with solubility approaching 50% in water, whereas OPA is hydrophobic in nature, with water solubility reaching a maximum at 5%, though it is highly soluble in alcohol and other organic solvents. OPA is at least five to eight times more active against mycobacteria than is glutaraldehyde when tested under the same conditions (295). Increased activity may be attributable to the hydrophobicity of the OPA molecule and its ability to penetrate the lipid cell wall layer. Phenol and phenolic compounds also exhibit better activity against the mycobacteria than more hydrophilic compounds. Several studies evaluating the role of the mycobacterial cell wall support the cell wall barrier hypothesis. The treatment of M. avium and M. tuberculosis with ethambutol, which acts by inhibiting a component of the mycobacterial cell wall, renders these organisms more susceptible to the activity of chlorhexidine and QACs (54).
Gram-negative organisms in general are less susceptible to germicides than are gram-positive bacteria. This is most likely due to the protective outer membrane in gram-negative organisms, which limits penetration of chemicals into the cell (25). It has been demonstrated that changes in the composition of the cell surface, especially the LPSs and lipids, can reduce or prevent the penetration of QACs into gram-negative cells (296). The cell wall of gram-positive bacteria may sometimes act as an important mechanism in the resistance of these bacteria to chemical agents, although this mechanism is not uniform across all species (297,298). In addition, the role of extracellular material in the protection of cells is further supported by the existence of mucoid and nonmucoid strains of S. aureus: The nonmucoid cells are more susceptible to several commonly used germicidal agents (299).
Studies on the adaptive resistance of Salmonella enterica and E. coli 0157 and cross-resistance to antimicrobial agents reported that E. coli 0157 exhibited a high level of resistance to triclosan after exposure to just two sublethal doses. This was accompanied by decreased susceptibility to a number of antibiotics, including chloramphenicol, erythromycin, tetracycline, trimethoprim, and imipenem. Cross-resistance in S. enterica was also demonstrated (300).
P. aeruginosa exhibits both intrinsic and acquired resistance. This organism is intrinsically resistant to many germicides and antibiotics. Within P. aeruginosa, a chromosomal DNA fragment conferring triclosan resistance has been isolated. The mexJK operon is responsible for the efflux mechanism required for triclosan effectiveness and, in conjunction with the outer membrane channel protein OprM, the mechanism required for erythromycin activity. Triclosan can select for mutants that are cross-resistant to antibiotics (301). Growth of Pseudomonas fluoresences in gradually higher concentrations of the QAC didecyl dimethylammonium chloride resulted in higher resistance and cross-resistance to several antibacterial agents. Microscopic observation of cells revealed slime formation as well as loss of flagella. Removal of the slime layer (a biofilm or a physical mechanism of resistance) resulted in loss of resistance (302).
Acquired Resistance
Acquired resistance to germicidal agents may occur through mutation or by acquisition of extrachromosomal DNA (plasmids or transposons). The role of plasmids in antibiotic resistance has been known for some time. In the past, it was thought that resistance to chemical germicides was not mediated through plasmids. A number of studies have indicated that plasmids do play a role in resistance to chemical germicides.
Acquired staphylococcal resistance to QACs has been reported extensively (303–306). Several genes involved in resistance to QACs have been isolated from strains of S. aureus and coagulase-negative staphylococcus (qac A, qacB, and qacC [smr]) (307–309) found in humans. Additionally, two genes, qacG and qacH, have been isolated from coagulase-negative staphylococcus identified by the food industry (303,304). A fifth gene encoding for resistance to QACs, qacJ, has been identified in several species of Staphylococcus isolated from horses (310). In addition, resistance of S. aureus to triclosan by a novel mechanism, sh-fabl derived from Staphylococcus haemolyticus, revealed the potential for horizontal gene transfer. This demonstrated that a biocide could spread resistance in a human pathogen (311).
Concern has been expressed over emerging cross-resistance or coresistance to widely used disinfectants and antibiotics (269). The qac genes have been found near antibiotic resistance genes for several antibiotics (gentamicin, kanamycin, tobramycin, trimethoprim, and penicillin) on several plasmids (pST6, pSK4, pSK410) and transposons (Tn552 and Tn4002) (305,307,312,313). In a survey of 522 clinical strains of S. aureus in Japanese hospitals, 32.6% of the MRSA strains and 7.5% of the methicillin-sensitive S. aureus strains also contained the qacA and qacB genes. The qacC (smr) gene was noted at a much lower frequency (314). Clinical isolates of staphylococci were resistant to a QAC, and benzalkonium chloride in 50% of cases (306). In those QAC-resistant strains, there was a high frequency of organisms also resistant to a range of antibiotics. The plasmid DNA of most isolates hybridized with one or more of the qac genes as well as blaZ (which codes for β-lactamase) and tetK (which codes for the tetracycline efflux pump). Approximately 34% of the plasmids had both the qac A/B and blaZ genes. One isolate had a multiresistant plasmid (pMS62) that contained the qacA/B, blaZ, and tetK genes.
Resistance to other antiseptics may also be found in some of these strains. Approximately 25% of the benzalkonium chloride–resistant strains also showed low-level resistance to the antiseptic chlorhexidine (314). A correlation between resistance to QACs and resistance to antibiotics in L. monocytogenes has also been reported (271). The genes responsible for this resistance can be plasmid-acquired (315) or chromosomal, similar to that found in S. aureus. The mdrL gene in L. monocytogenes forms a protein that is similar to efflux mechanism proteins in B. subtilis (316). Originally, mdrL was reported to be a chromosomal gene (317), that is, intrinsic resistance. However, further work demonstrated that it may be chromosomal or plasmid-acquired. Chlorhexidine resistance in K. pneumoniae has been linked to the cepA gene (associated with the production of cephalosporinase). Transformation of the cepA to the E. coli XLOLR resulted in chlorhexidine sensitivity, and retransformation resulted in chlorhexidine-sensitive K. pneumonia. It is hypothesized that the gene product CepA may act as a cation efflux pump. The incorporation of triclosan in many household products may contribute to the problem of increased antibiotic resistance (270), yet evidence collected in environmental samples (318,319) and human skin flora (320) does not support this supposition. The accumulated data suggest that resistance to antiseptics and disinfectants may impart antibiotic resistance in some instances (321,322). Table 12.21 reviews genes that may be responsible for shared resistance to both antibiotics and disinfectants. Consideration of these important potential interactions must be addressed when assessing isolates from clinical human samples.
Anticipated Role of Disinfectants and Antiseptics in the Health Care Environment
Hospital-acquired or nosocomial infections are the major cause of morbidity and mortality in patients contributing to health care costs and prolonged lengths of hospitalization (1). As medical practice extends beyond inpatient hospital settings, with an increasing outpatient presence, including at-home care, the more inclusive phrase “health care–acquired infection” should be used (323). The IHI (1) has focused the medical community on the need to control and prevent such infections. This chapter has addressed the commonly used disinfectants and antiseptics; however, many issues remain regarding health care–acquired infections and the health care environment (1,268,322,323).
■ There are continued concerns regarding multiresistant microorganisms in the environment (e.g., MRSA and VRE).
■ Data continue to reveal inadequate handwashing by health care workers and the contribution to contamination of the health care environment.
■ Concerns remain regarding prior hospital or institutional room utilization by an infected patient and the risk of contamination to a new room occupant.
■ The mandate to identify and incorporate updated methods of terminal cleaning of hospital or institutional rooms, such as ultraviolet light treatment and vaporized hydrogen peroxide generators (211,324,325) continues.
■ There is an ongoing need to perform continuous surveillance of the health care environment to assess microorganism burden and changing patterns of resistance.
■ Perform frequent updates and education for health care and environmental health services, as well as patients and visitors, about the risk of health-care acquired infections.
Innovative approaches supported by laboratory methods using antiseptics and disinfectants in creative ways, such as vaporization and ingredient combinations, will further minimize the risk of microorganism transmission. These tactics will enhance health care institutions’ efforts to achieve The Joint Commission’s 2013 National Patient Safety Goals, including improved hand hygiene and decrease/reduce surgical site infections, catheter-associated line infections, and urinary tract infections (326). Fewer health care–acquired infections will ultimately reduce morbidity and mortality, coming closer to achieving the crucial goals set by the IHI. It is likely that these goals will be incorporated into the Centers for Medicare & Medicaid Services (CMS) “value-based purchasing” initiatives that will tie diagnosis-related group (DRG) and medical reimbursement schema to performance in the coming years.
REFERENCES
1. Institute for Healthcare Improvement. 5 million lives campaign. 2008. http://www.ihi.org/offerings/Initiatives/PastStrategicInitiatives/5MillionLivesCampaign. Accessed November 15, 2012.
2. Ducel G, Fabry J, Nicolle L, eds. World Health Organization Prevention of hospital-acquired infections. A practical guide. 2nd ed. Geneva: World Health Organization, 2002:1–3.
3. Occupational Safety and Health Administration. U.S. Department of Labor compliance with the OSHA bloodborne pathogens standard 29 CFR 1910.1030. http://www.osha.gov/pls/oshaweb/owadisp. Accessed November 15, 2012.
4. Rutala W, Weber D; Healthcare Infection Control Practices Advisory Committee, Centers for Disease Control and Prevention. Guideline for disinfection and sterilization in healthcare facilities 2008. http://www.cdc.gov/hicpac/Disinfection_Sterilization/table. Accessed November 15, 2012.
5. Spaulding EH. Chemical disinfection of medical and surgical disinfection of medical and surgical materials. In: Lawrence CA, Block SS, eds. Disinfection, sterilization and preservation. Philadelphia: Lea & Febiger, 1968:517–531.
6. Garner JS, Favero MS. Guidelines for handwashing and hospital environmental control, 1985. Am J Infect Control 1986;14:110–126.
7. Boyce JM. Methicillin-resistant Staphylococcus aureus in hospitals and long-term care facilities: microbiology, epidemiology, and preventive measures. Infect Control Hosp Epidemiol 1992;13:725–737.
8. Boyce JM, Jackson MM, Pugliese G, et al. Methicillin-resistant Staphylococcus aureus (MRSA): a briefing for acute care hospitals and nursing facilities. Infect Control Hosp Epidemiol 1994;15:105–115.
9. Mulligan ME, Murray-Leisure KA, Ribner BS, et al. Methicillin-resistant Staphylococcus aureus: a consensus review of the microbiology, pathogenesis, and epidemiology with implications for prevention and management. Am J Med 1993;94:311–328.
10. Weber DJ, Rutala WA. Environmental issues and nosocomial infections. In: Wenzel RP, ed. Prevention and control of nosocomial infections. 3rd ed. Baltimore: Lippincott Williams & Wilkins, 1997:491–514.
11. Kim KH, Fekety R, Batts DH, et al. Isolation of Clostridium difficile from the environment and contacts of patients with antibiotic-associated colitis. J Infect Dis 1981;143:42–50.
12. Larson HE, Barclay FE, Honour P, et al. Epidemiology of Clostridium difficile in infants. J Infect Dis 1982;146:727–733.
13. Malamou-Ladas H, O’Farrell S, Nash JQ, et al. Isolation of Clostridium difficile in hospitalized children: a prospective study. Acta Paediatr Scand 1990;79:292–299.
14. Kaatz GW, Gitlin SD, Schaberg DR, et al. Acquisition of Clostridium difficile from the hospital environment. Am J Epidemiol 1988;127:1289–1294.
15. Boyce JM. Scientific basis for hand washing with alcohol and other waterless antiseptic agents. In: Rutala WA, ed. Disinfection, sterilization and antisepsis: principles and practices in healthcare facilities. Washington, DC: Association for Professionals in Infection Control and Epidemiology, 2001:140–150.
16. Longtin Y, Sax H, Allegranzi B, et al. Hand hygiene. N Engl J Med 2011;364:13–17.
17. Sax H, Allegranzi B, Chraiti MN, et al. The WHO hand hygiene observation method. Am J Infect Control 2009;37:827–834.
18. Rotter ML. Hand washing, hand disinfection and skin disinfection. In: Wenzel RP, ed. Prevention and control of nosocomial infections. 3rd ed. Baltimore: Lippincott Williams & Wilkins, 1997:691–709.
19. Ascenzi J, Favero M. Disinfectants and antiseptics: modes of action, mechanisms of resistance and testing. In: Lorian V, ed. Antibiotics in laboratory medicine. 5th ed. Philadelphia: Lippincott Williams & Wilkins, 2005:615–653.
20. Williams G, Denyer S, Hosein I, et al. The development of a new three-step protocol to determine the efficacy of disinfectant wipes on surfaces contaminated with S. aureus. J Hosp Infect 2007;67:329–335.
21. Siani H, Cooper C, Maillard JY. Efficacy of “sporicidal” wipes against Clostridium difficile. Am J Infect Control 2011;39:212–218.
22. McDonnell G, Burke P. Disinfection: is it time to reconsider Spaulding? J Hosp Infect 2011;78:163–170.
23. European Union. Regulation (EU) No 528/2012 of the European Parliament and of the Council of 22 May 2012 concerning the making available on the market and use of biocidal products. Official J of EU 2012;1–123. http://eur-lex.europa.eu/LexUriServ/LexUriServ.do. Accessed November 15, 2012.
24. Sharma BK, Shamim A, Lalia A, et al. Chemical agents as disinfectants and antiseptics: a review. The Global J of Pharmaceutical Res 2012;1:795–803.
25. McDonnell G, Russell AD. Antiseptics and disinfectants: activity, action, and resistance. Clin Microbiol Rev 1999;12:147–179.
26. Mazzola P, Jozala A, Lencastre Novaes L, et al. Minimal inhibitory concentration (MIC) determination of disinfectant and/or sterilizing agents. Braz J Pharm Sci 2009;45:241–248.
27. Mazzola P, Jozala A, Lencastre Novaes L, et al. Choice of sterilizing/disinfecting agent—determination of the Decimal Reduction Time (D-Value). Braz J Pharm Sci 2009;45:701–718.
28. McDonnell GE. Antisepsis, disinfection and sterilization. Types, action and resistance. Washington, DC: ASM Press, 2007.
29. Sattar SA, Springthorpe VS. New methods for efficacy testing of disinfectants and antiseptics. In: Rutala WA, ed. Disinfection, sterilization and antisepsis: principles and practices in healthcare facilities. Washington, DC: Association for Professionals in Infection Control and Epidemiology, 2001:173–186.
30. Herruzo R, Vizcaino MJ, Herruzo I. In vitro-in vivo sequence studies as a method of selecting the most efficacious alcohol-based solution for hygienic hand disinfection. Clin Microbiol Infect 2010;16:518–523.
31. Ramirez-Arcos S, Goldman M. Skin disinfection methods: prospective evaluation and postimplementation results. Transfusion 2010;50:59–64.
32. Rotter ML, Koller W, Wewalka G. Povidone-iodine and chlorhexidine gluconate–containing detergents for disinfection of hands. J Hosp Infect 1980;1:149–158.
33. Hobson DW, Bolsen K. Methods of testing oral and topical antiseptics and antimicrobials. In: Block SS, ed. Disinfection, sterilization and preservation. 5th ed. Philadelphia: Lippincott Williams & Wilkins, 2001:1329–1359.
34. Sattar SA, Springthorpe VS. Methods under development for evaluating the antimicrobial activity of chemical germicides. In: Rutala WA, ed. Chemical germicides in health-care. 3rd ed. Morin Heights, Canada: Polyscience and Association for Professionals in Infection Control and Epidemiology, 1994:237–254.
35. Sattar SA. Microbiocidal: testing of germicides: an update. In: Rutala WA, ed. Disinfection, sterilization, and antisepsis in healthcare. Washington, DC: Association for Professionals in Infection Control and Epidemiology, 1998:227–240.
36. Miner NA, Mulberry GK, Starks AN, et al. Identification of possible artifacts in the Association of Official Analytical Chemists sporicidal test. Appl Environ Microbiol 1995;61:1658–1660.
37. Ascenzi JM, Ezzell JM, Wendt TM. A more specific method for measurement of tuberculocidal activity of disinfectants. Appl Environ Microbiol 1987;53:2189–2192.
38. Springthorpe VS, Sattar SA. Application of a quantitative carrier test to evaluate microbiocides against mycobacteria. J AOAC Int 2007;90:817–824.
39. Steinmann J. Surrogate viruses for testing virucidal efficacy of chemical disinfectants. J Hosp Infect 2004;56:549–554.
40. Association of Official Analytical Chemists International. Disinfectants: sporicidal activity of disinfectants. Washington, DC: Association of Official Analytical Chemists, Official Methods of Analysis, 1998.
41. Humphreys PN. Testing standards for sporicides. J Hosp Infect 2011;77:193–198.
42. Beyth N, Redlich M, Harari D, et al. Effect of sustained-release chlorhexidine varnish of Streptococcus mutans and Actinomyces viscosus in orthodontic patients. Am J Orthod Dentofacial Orthop 2003;123:345–348.
43. Denton GW. Chlorhexidine. In: Block SS, ed. Disinfection, sterilization and preservation. 5th ed. Philadelphia: Lippincott Williams & Wilkins, 2001:321–336.
44. Davies GE, Field BS. Action of biguanides, phenols, and detergents on E. coli and its spheroplasts. J Appl Bacteriol 1969;32:233–243.
45. Hugo WB, Longworth AR. Some aspects of mode of action of chlorhexidine. J Pharm Pharmacol 1964;16:655–662.
46. Hugo WB, Longworth AR. Cytological aspects of the mode of action of chlorhexidine. J Pharm Pharmacol 1965;17:28–32.
47. Hugo WB, Longworth AR. The effect of chlorhexidine on the electrophoretic mobility, cytoplasmic constituents, dehydrogenase activity and cell walls of E. coli and S. aureus. J Pharm Pharmacol 1966;18:569–578.
48. Fitzgerald KA, Davies A, Russell AD. Uptake of 14C-chlorhexidine diacetate to Escherichia coli and Pseudomonas aeruginosa and its release by azolectin. FEMS Microbiol Lett 1989;60:327–332.
49. Hiom SJ, Furr JR, Russell AD. Effects of chlorhexidine diacetate on Candida albicans, C. glabrata, and Saccharomyces cerevisiae. J Appl Bacteriol 1992;72:335–340.
50. Tattawasart U, Maillard J-Y, Furr JR, et al. Outer membrane changes in Pseudomonas stutzeri resistant to chlorhexidine diacetate and cetylpyridinium chloride. Int J Antimicrob Agents 2000;16:233–238.
51. Tattawasart U, Hann AC, Maillard JY, et al. Cytological changes in chlorhexidine-resistant isolates of Pseudomonas stutzeri. J Antimicrob Chemother 2000;45:145–152.
52. Ranganathan NS. Chlorhexidine. In: Ascenzi JM, ed. Handbook of disinfectants and antiseptics. New York: Marcel Dekker, 1996:235–264.
53. Sheppard FC, Mason DJ, Bloomfield SF, et al. Flow cytometric analysis of chlorhexidine action. FEMS Mircobiol Lett 1997;2:283–288.
54. Broadley SJ, Jenkins PA, Furr JR, et al. Potentiation of effects of chlorhexidine diacetate and cetylpyridinium chloride on mycobacteria by ethambutol. J Med Microbiol 1995;43:458–460.
55. Walters TH, Furr JR, Russell AD. Antifungal action of chlorhexidine. Microbios 1983;38:195–204.
56. Bobichon H, Bouchet P. Action of chlorhexidine on budding of Candida albicans: scanning and transmission electron microscopic study. Mycopathologia 1987;100:27–35.
57. Shaker LA, Russell AD, Furr JR. Aspects of the action of chlorhexidine on bacterial spores. Int J Pharm 1986;34:51–56.
58. Holloway PM, Bucknall RA, Denton GW. The effect of sub-lethal concentrations of chlorhexidine on bacterial pathogenicity. J Hosp Infect 1986;8:39–46.
59. Montefiori DC, Robinson WE, Modliszewski A, et al. Effective inactivation of human immunodeficiency virus with chlorhexidine antiseptics containing detergents and alcohol. J Hosp Infect 1990;15:279–282.
60. Harbison MA, Hammer SM. Inactivation of human immunodeficiency virus by Betadine and chlorhexidine. J AIDS 1989;2:16–20.
61. Kaiser N, Klein D, Karanja P, et al. Inactivation of chlorhexidine gluconate on skin by incompatible alcohol hand sanitizing gels. J Infect Control 2009;37:569–573.
62. Peterson AF, Rosenberg A, Alatary SD. Comparative evaluation of surgical scrub preparations. Surg Gynecol Obstet 1978;146:63–65.
63. Wade JJ, Casewell MW. The evaluation of residual antimicrobial activity on hands and its clinical relevance. J Hosp Infect 1991;18(Suppl B):23–28.
64. Mulberry G, Snyder AT, Heilman J, et al. Evaluation of a waterless, scrubless chlorhexidine gluconate/ethanol scrub for antimicrobial efficacy. Am J Infect Control 2001;29:377–382.
65. Larson EL, Laughon BE. Comparison of four antiseptic products containing chlorhexidine gluconate. Antimicrob Agents Chemother 1987;31:1572–1574.
66. Hibbard JS, Mulberry GK, Brady AR. A clinical study comparing the skin antisepsis and safety of ChloraPrep, 70% isopropyl alcohol, and 2% aqueous chlorhexidine. J Infus Nurs 2002;25:244–249.
67. Wilcox MH, Hall J, Gill AB, et al. Effectiveness of topical chlorhexidine powder as an alternative to hexachlorophene for the control of Staphylococcus aureus in neonates. J Hosp Infect 2004;56:156–159.
68. Maki DG, Ringer M, Alvarado CJ. Prospective randomized trial of povidone-iodine, alcohol, and chlorhexidine for prevention of infection associated with central venous and arterial catheters. Lancet 1991;228:339–343.
69. Clemence MA, Walker D, Farr BM. Central venous catheter practices: results of a survey. Am J Infect Control 1995;23:5–12.
70. Mimoz O, Pieroni L, Lawerence C, et al. Prospective randomized trial of two antiseptic solutions for prevention of central venous or arterial catheter colonization and infection in intensive care unit patients. Crit Care Med 1996;24:1818–1823.
71. Rubinson L, Diette GB. Best practices for insertion of central venous catheters in intensive-care units to prevent catheter-related bloodstream infections. J Lab Clin Med 2004;143:5–13.
72. Walder B, Pittet D, Tramer MR. Prevention of bloodstream infections with central venous catheters treated with anti-infective agents depends on catheter type and insertion time: evidence from a meta-analysis. Infect Control Hosp Epidemiol 2002;23:748–756.
73. Heard O, Wagle M, Vijayakumar E, et al. Influence of triple lumen catheter venous catheters coated with chlorhexidine and silver sulfadiazine on the incidence of catheter-related bacteremia. Arch Intern Med 1998;158:81–87.
74. Magnusson I. Local delivery of antimicrobial agents for the treatment of periodontitis. Compend Contin Educ Dent 1998;19:953–956.
75. Tomás I, García-Caballero L, Cousido MC, et al. Evaluation of chlorhexidine substantivity on salivary flora by epifluorescence microscopy. Oral Dis 2009;15:428–433.
76. Turner LA, McCombs GB, Hynes WL, et al. A novel approach to controlling bacterial contamination on toothbrushes: chlorhexidine coating. Int J Dent Hyg 2009;7:241–245.
77. Chavan SD, Shetty NL, Kanuri M. Comparative evaluation of garlic extract mouthwash and chlorhexidine mouthwash on salivary Streptococcus mutans count—an in vitro study. Oral Health Prev Dent 2010;8:369–374.
78. Eick S, Goltz S, Nietzsche S, et al. The efficacy of chlorhexidine digluconate containing formulations and other mouth rinses against periodontopathogenic microorganisms. Quintessence Int 2011;42:687–700.
79. Labeau SO, Van de Vyver K, Brusselaers N, et al. Prevention of ventilator-associated pneumonia with oral antiseptics: a systematic review and meta-analysis. Lancet Infect Dis 2011;11:845–854.
80. Piñeiro A, Tomás I, Blanco J, et al. Bacteremia following dental implant placement. Clin Oral Implants Res 2010;21:913–918.
81. Stokes T, Richey R, Wrayon D. Prophylaxis against endocarditis: summary of NICE guidelines. Heart 2008;94:930–931.
82. Favero MS, Bond WW. Chemical disinfection of medical and surgical materials. In: Block SS, ed. Disinfection, sterilization and preservation. 5th ed. Philadelphia: Lippincott Williams & Wilkins, 2001:881–918.
83. Berkelmann RI, Holland BW, Anderson RI. Increased bactericidal activity of dilute preparations of povidone-iodine solution. J Clin Microbiol 1982;15:635–639.
84. Gottardi W. Iodine and iodine compounds. In: Block SS, ed. Disinfection, sterilization and preservation. 5th ed. Philadelphia: Lippincott Williams & Wilkins, 2001:159–183.
85. Weber D, Rutala W, Sickert-Bennett E. Outbreaks associated with contaminants of antiseptics and disinfectants. Antimicrob Agents Chemother. 2007;51:4217–4224.
86. Craven DE, Moody B, Connolly BS, et al. Pseudobacteremia caused by povidone-iodine solution contaminated with Pseudomonas aeruginosa. N Eng J Med 1981;305:621–623.
87. Gottardi W, Puritscher M. Degerming experiments with aqueous povidone-iodine containing disinfecting solutions: influence of the concentration of free iodine on the bactericidal reaction against Staphylococcus aureus. Zentralbl Bakteriol 1986;182:372–380.
88. Bloomfield SA. Chlorine and iodine formulations. In: Ascenzi JM, ed. Handbook of disinfectants and antiseptics. New York: Marcel Dekker, 1996:133–158.
89. Wei MK, Wu QP, Huang Q, et al. Plasma membrane damage to Candida albicans caused by chlorine dioxide (ClO2). Lett Appl Microbiol 2008;47:67–73.
90. Gershenfeld L, Witlin B. Iodine solution as a sporicidal agent. J Am Pharm Assoc 1952;41:451–452.
91. Bartlett PG, Schmidt W. Disinfectant iodine complexes as germicides. J Appl Microbiol 1957;5:355–359.
92. Sykes G. The sporicidal properties of chemical disinfectants. J Appl Bacteriol 1970;33:147–156.
93. Russell AD. Chemical sporicidal and sporostatic agents. In: Block SS, ed. Disinfection, sterilization and preservation. 5th ed. Philadelphia: Lippincott Williams & Wilkins, 2001:529–541.
94. Pyle BH, McFeters GA. Iodine sensitivity of bacteria isolated from iodinated water systems. Can J Microbiol 1989;35:520–523.
95. Cargill KL, Pyle BH, Sauer RL, et al. Effects of culture conditions and biofilm formation on iodine susceptibility of Legionella pneumophila. Can J Microbiol 1992;38:423–429.
96. Gershenfeld L, Flagg W, Witlin B. Iodine as a tuberculocidal agent. Mil Surg 1954;114:172–183.
97. Nelson KE, Larson PA, Schraufnagel DE, et al. Transmission of tuberculosis by flexible fiberbronchoscopes. Am Rev Respir Dis 1983;127:97–100.
98. Best M, Sattar SA, Springthorpe VS, et al. Efficacies of selected disinfectants against Mycobacterium tuberculosis. J Clin Microbiol 1990;28:2234–2239.
99. Best M, Sattar SA, Springthorpe VS, et al. Comparative mycobactericidal efficacy of chemical disinfectants in suspension and carrier tests. Appl Environ Microbiol 1988;54:2856–2858.
100. Arzmi MH, Abdul R, Yusoff M, et al. Effect of phenotype switching on the biological properties and susceptibility to chlorhexidine in Candida krusei. FEMS Yeast Res 2012;12:351–358.
101. Berg G, Chang SL, Harris EK. Devitalization of microorganisms by iodine. Virology 1964;22:469–481.
102. Gershenfeld L. Iodine as a virucidal agent. J Am Pharm Assoc 1955;44:177–182.
103. Prince HN, Prince DL. Principles of viral control and transmission. In: Block SS, ed. Disinfection, sterilization and preservation. 5th ed. Philadelphia: Lippincott Williams & Wilkins, 2001:543–571.
104. Sauerbrei A, Wutzler P. Virucidal efficacy of povidone iodine containing disinfectants. Letters Applied Microbiol 2010;51:158–163.
105. Thraenhart O, Jursch C. Measures for disinfection and control of viral hepatitis. In: Block SS, ed. Disinfection, sterilization and preservation. 5th ed. Philadelphia: Lippincott Williams & Wilkins, 2001:585–615.
106. Crabtree TD, Pelletier SJ, Pruett TL. Surgical antisepsis. In: Block SS, ed. Disinfection, sterilization and preservation. 5th ed. Philadelphia: Lippincott Williams & Wilkins, 2001:919–934.
107. Ayliffe GA, Babb JR, Davies JG, et al. Hand disinfection: a comparison of various agents in laboratory and ward studies. J Hosp Infect 1988;11:226–243.
108. Lowbury EJ, Lilly HA. The effect of blood on disinfection of surgeons’ hands. Br Surg J 1974;61:19–21.
109. Ally R, Maibach H. Comparative evaluation of chlorhexidine gluconate (Hibiclens) and povidone-iodine (E-Z Scrub) sponge/brushes for presurgical hand scrubbing. Curr Ther Res 1983;34:740–745.
110. Rotter ML. Alcohols for antisepsis of hands and skin. In: Ascenzi JM, ed. Handbook of disinfectants and antiseptics. New York: Marcel Dekker, 1996:177–233.
111. Kamm O. The relation between structure and physiological action of the alcohols. J Am Pharm Assoc 1921;10:87–92.
112. Sykes G. Disinfection and sterilization. 2nd ed. London: E & FN Spon Ltd, 1965:362–376.
113. Soberheim G. Alkohol als Disinfektionsmittel. Schweiz Med Wochenschr 1943;73:1280–1333.
114. Sykes G. Influence of germicides on dehydrogenase of Bacterium coli: succinic acid dehydrogenase of Bacterium coli. J Hyg 1939;59:463–469.
115. Kirschhoff H. Wirkungmechanismem chemischer Desinfektionsmittel. I. Allgemeiner Reaktionsablauf. Gesundheitwes Desinfekt 1974;66:125–130.
116. Pulvertaft RJV, Lumb GD. Bacterial lysis and antiseptics. J Hyg (London) 1948;46:62–64.
117. Razin S, Argaman M. Lysis of Mycoplasma, bacterial protoplasts, spheroplasts and L-forms by various agents. J Gen Microbiol 1963;30:155–172.
118. Harrington C, Walker H. The germicidal activity of alcohol. Boston Med Surg J 1903;148:548–552.
119. Coulthard CE, Sykes G. The germicidal effect of alcohol with special reference to its action on bacterial spores. Pharm J 1936;137:79–81.
120. Pohle WD, Stuart LS. The germicidal action of cleaning agents: a study of a modification of Price’s procedure. J Infect Dis 1940; 67:275–281.
121. Sakuragi T, Yanagisawa K, Dan K. Bactericidal activity of skin disinfectants on methicillin-resistant Staphylococcus aureus. Anesth Analg 1995;81:555–558.
122. Kampf G, Jarosch R, Ruden H. Limited effectiveness of chlorhexidine based hand disinfectants against methicillin resistant Staphylococcus aureus (MRSA). J Hosp Infect 1998;38:297–303.
123. Kampf G, Hofer M, Wendt C. Efficacy of hand disinfectants against vancomycin resistant enterococci. J Hosp Infect 1999;42:143–150.
124. Morton HW. Relationship of concentration and germicidal efficacy of ethyl alcohol. Ann NY Acad Sci 1950;532:191–196.
125. Ali Y, Dolan MJ, Fendler EJ, et al. Alcohols. In: Block SS, ed. Disinfection, sterilization and preservation. 5th ed. Philadelphia: Lippincott Williams & Wilkins, 2001:229–254.
126. Woo PCY, Leung K-W, Wong SSY, et al. Relatively alcohol-resistant Mycobacteria are emerging pathogens in patients receiving acupuncture treatment. J Clin Microbiol 2002;40;1219–1224.
127. Gordon MH. Studies on viruses of vaccinia and variola. Privy Council Medical Research Council, Special Reports 1925; Series 98.
128. Kuwert EK, Thraendhardt O. Theoretische, Methodische und praktische Probleme der Virusdesinfektion in der Hummanmedizin. Immun Infekt 1977;4:125–130.
129. Groupe V, Engle CG, Gaffney PE, et al. Virucidal activity of representative anti-infective agents against influenza A and vaccinia viruses. Appl Microbiol 1955;3:333–339.
130. Klein M, Deforest A. The inactivation of virus by germicides. Proc Chem Spec Manuf 1963:116–118.
131. Kewitsch A, Weuffen W. Wirkung chemischer Desinfektionsmittel gegenuber Influenza-Vacciniaund Poliomyelitisvirus. Med Welt 1966;17:76–81.
132. Mbithi JN, Springthorpe VS, Sattar SA. Chemical disinfection of hepatitis A virus on environmental surfaces. Appl Env Microbiol 1990;56:3601–3604.
133. Bond WW, Favero MS, Petersen NJ, et al. Inactivation of hepatitis B virus by intermediate-to-high level disinfectant chemicals. J Clin Microbiol 1983;18:535–538.
134. Kobayashi H, Tsuzuki M, Koshimizu K, et al. Susceptibility of hepatitis B virus to disinfectants or heat. J Clin Microbiol 1984;20:214–216.
135. Ehrenkranz HN, Alfonso BC. Failure of hand washing to prevent hand transfer of patient bacteria to urethral catheters. Infect Control Hosp Epidemiol 1991;12:654–658.
136. Handley JO, Mika LA, Gwaltney JM. Evaluation of virucidal compounds for inactivation of rhinovirus on hands. Antimicrob Agents Chemother 1978;14:690–694.
137. Savolainen-Kopra C, Korpela T, Simonen-Tikka ML, et al. Single treatment with ethanol hand rub is ineffective against human rhinovirus—hand washing with soap and water removes the virus efficiently. J Med Virol 2012;84:543–547.
138. Grayson ML, Melvani S, Druce J, et al. Efficacy of soap and water and alcohol-based hand-rub preparations against live H1N1 influenza virus on the hands of human volunteers. Clin Infect Dis 2009;48:285–291.
139. Pepper RE, Lieberman ER. Dialdehyde alcoholic sporicidal composition. US Patent 3,016,328. January 9, 1962.
140. Rehn D, Nolte H. Zur antimikrobiellen wirksamkeit substituierter aromatischer aldehyde und alkohole. Zentrabl Bakteriol Hyg Abt I Orig B 1979;168:506–516.
141. Hopewood D, Allen CR, McCabe C. The reactions between glutaraldehyde and various proteins: an investigation of their kinetics. Histochem J 1970; 2:137–150.
142. Ellar DJ, Munoz E, Salton MRJ. The effect of low concentrations of glutaraldehyde on Micrococcus lysodeikticus membranes. Biochim Biophys Acta 1971;225:140–150.
143. Chambon M, Jallat-Archimbaud C, Bailly JL, et al. Comparative sensitivities of Sabin and Mahoney poliovirus type 1 prototype strains and two recent isolates to low concentrations of glutaraldehyde. Appl Environ Microbiol 1997;63:3199–3204.
144. Howard CR, Dixon JL, Young P, et al. Chemical inactivation of hepatitis B virus: the effect of disinfectants on virus associated DNA polymerase activity, morphology and infectivity. J Virol Methods 1983;7:135–148.
145. Fraenkel-Conrat H, Cooper M, Olcott HS. The reaction of formaldehyde with proteins. J Am Chem Soc 1945;67:950–954.
146. Staehlin M. Reaction of tobacco mosaic virus nucleic acid with formaldehyde. Biochim Biophys Acta 1958;29:410–417.
147. Simoes M, Simoes LC, Cleto S, et al. Antimicrobial mechanisms of ortho-phthalaldehyde action. J Basic Microbiol 2007;47:230–242.
148. Gorman SP, Scott EM. Transport capacity, alkaline phosphatase activity and protein content of glutaraldehyde-treated cell forms of Escherichia coli. Microbios 1977;19:205–212.
149. Fraud S, Hann AC, Maillard J-Y, et al. Effect of ortho-phthalaldehyde, glutaraldehyde and chlorhexidine diacetate on Mycobacterium chelonae and Mycobacterium abscessus strains with modified permeability. J Antimicrob Chemother 2003;51:575–584.
150. McErlean EP, Gorman SP, Scott EM. Physical and chemical resistance of ion-exchange and coat defective spores of Bacillus subtilis. J Pharm Pharmacol 1980;32:32P.
151. Gorman SP, Scott EM, Hutchinson EP. Interaction of Bacillus subtilis spore protoplast, cortex, ion-exchange and coatless forms with glutaraldehyde. J Appl Bacteriol 1984;56:95–102.
152. Gorman SP, Hutchison EP, Scott EM, et al. Death, injury and revival of chemically treated Bacillus subtilis spores. J Appl Bacteriol 1983;54:91–99.
153. Stonehill AA, Krop S, Borick PM. Buffered glutaraldehyde, a new chemical sterilizing solution. Am J Hosp Pharm 1963;20:458–465.
154. Rubo SD, Gardner JF, Webb RL. Biological activities of glutaraldehyde and related compounds. J Appl Bacteriol 1967;30:78–87.
155. Borick PM. Chemical sterilizers. Adv Appl Microbiol 1968;10:291–312.
156. Akamatsu T, Tabata K, Hironga M, et al. Transmission of Helicobacter pylori infection via flexible fiberoptic endoscopy. Am J Infect Control 1996;24:396–401.
157. Chan-Myers H, Roberts C, Ascenzi J. Virucidal activity of o-phthalaldehyde solutions against drug resistant bacteria. Abstracts of American Society for Microbiology Meeting 2001; Q64:595
158. Fraud S, Hann AC, Maillard J-Y, et al. Comparison of the mycobacteriocidal activity of ortho-phthalaldehyde, glutaraldehyde and other dialdehydes by a quantitative suspension test. J Hosp Infect 2001;48:214–221.
159. Collins FM, Montalbine V. Mycobacteriocidal activity of glutaraldehyde solutions. J Clin Microbiol 1976;4:408–412.
160. Collins FM. Bactericidal activity of alkaline glutaraldehyde solution against a number of atypical mycobacterial species. J Appl Bacteriol 1986;61:247–251.
161. Hernandez A, Martró E, Matas L, et al. In-vitro evaluation of Persafe compared with 2% alkaline glutaraldehyde against Mycobacterium spp. J Hosp Infect 2003;54:52–56.
162. Hernandez A, Martró E, Puzo C, et al. In-use evaluation of Persafe compared with Cidex in fiberoptic bronchoscope disinfection. J Hosp Infect 2003;54:46–51.
163. Carson LA, Favero MS, Bond WW, et al. Factors affecting comparative resistance of naturally occurring and subcultured Pseudomonas aeruginosa to disinfectants. Appl Microbiol Biotechnol 1972;23:863–869.
164. Carson LA, Petersen NJ, Favero MS, et al. Growth characteristics of atypical mycobacteria in water and their comparative resistance to disinfectants. Appl Env Microbiol 1978;36:839–846.
165. Sagripanti J-L, Bonafacino A. Comparative sporicidal effects of liquid chemical agents. Appl Environ Microbiol 1996;62:545–551.
166. Gorman SP, Scott EM. Effect of alkalination of the bacterial cell and glutaraldehyde molecule. Microbio Lett 1977;6:39–44.
167. Bartlett JG, Onderdonk AB, Cisneros RL, et al. Clindamycin-associated colitis due to toxin-producing species of Clostridium in hamsters. J Infect Dis 1977;136:701–705.
168. Bartlett JG, Chang TW, Gurwith M, et al. Antibiotic-associated pseudo-membranous colitis due to toxin producing clostridia. N Engl J Med 1978;298:531.
169. Larson HE, Price AB, Honour P, et al. Clostridium difficile and the etiology of pseudomembranous colitis. Lancet 1978;1:1063–1066.
170. Teasley DG, Gerding DN, Olson MM. Prospective randomized trial of metronidazole versus vancomycin for Clostridium difficile–associated diarrhea and colitis. Lancet 1983;2:1043–1046.
171. Dyas A, Das BC. The activity of glutaraldehyde against Clostridium difficile. J Hosp Infect 1985;6:41–45.
172. Hughes CE, Gerhard RL, Petersen LR, et al. Efficacy of routine fiberoptic endoscope cleaning and disinfection for killing Clostridium difficile. Gastrointest Endosc 1986;32:7–9.
173. Oie S, Kamiya A. Sporicidal activity of aldehyde disinfectants. Env Infect 2003;18:1–9.
174. Gorman SP, Scott EM. A quantitative evaluation of the antifungal activity of glutaraldehyde. J Pharm Pharmacol 1977;43:83–89.
175. Terleckyj B, Axler DA. Quantitative neutralization assay of fungicidal properties of disinfectants. Antimicrob Agents Chemother 1987;31:794–798.
176. Scott EM, Gorman SP. Glutaraldehyde. In: Block SS, ed. Disinfection, sterilization and preservation. 5th ed. Philadelphia: Lippincott Williams & Wilkins, 2001:361–381.
177. Dabrowa N, Landau JW, Newcomer VD. Antifungal activity of glutaraldehyde in vitro. Arch Dermatol 1972;105:555–557.
178. Suringa DWR. Treatment of superficial onychomycoses with topically applied glutaraldehyde. Arch Dermatol 1970;102:163–167.
179. Isenberg HD, Giugliano ER, France K, et al. Evaluation of three disinfectants after in-use stress. J Hosp Infect 1988;11:278–285.
180. Narang HK, Codd AA. Action of commonly used disinfectants against enteroviruses. J Hosp Infect 1983;4:209–212.
181. Saitanu K, Lund E. Inactivation of enterovirus by glutaraldehyde. Appl Microbiol 1975;29:571–574.
182. Cunliffe HR, Blackwell JH, Walker JS. Glutaraldehyde inactivation of exotic animal viruses in swine heart tissue. Appl Env Microbiol 1979; 37:1044–1046.
183. Spire B, Montagnier L, Barre-Sinoussi F, et al. Inactivation of lymphadenopathy associated virus by chemical disinfectants. Lancet 1984;2:899–901.
184. Hanson PJV, Gor D, Jefferies DJ, et al. Chemical inactivation of HIV on surfaces. Br Med J 1989;298:862–864.
185. Hanson PJV, Gor D, Clarke JR, et al. Contamination of endoscopes used in patients with AIDS. Lancet 1989;2:86–88.
186. Hanson PJV, Gor D, Jefferies DJ, et al. Elimination if high titer HIV from fiberoptic endoscopes. Gut 1990; 31:657–660.
187. Druce JD, Jardine D, Locarnini SA, et al. Susceptibility of HIV to inactivation by disinfectants and ultraviolet light. J Hosp Infect 1995;30:167–180.
188. Blackwell JH, Chen JHS. Effects of various germicidal chemicals on HEp 2 cell culture and herpes simplex virus. J Assoc Off Anal Chem 1970; 53:1229–1236.
189. Prince DL, Prince RN, Prince HN. Inactivation of human immunodeficiency virus type 1 and herpes simplex virus type 2 by commercial hospital disinfection. Chem Times Trends 1990:13–16.
190. Sable FL, Hellman A, McDade J. Glutaraldehyde inactivation of virus in tissue. Appl Microbiol 1969;17:645–646.
191. Seefeld U, Bansky G, Jaeger M, et al. Prevention of hepatitis B virus transmission by gastrointestinal fiberscope: successful disinfection with an aldehyde liquid. Endoscopy 1981;13:238–239.
192. Adler-Storthz K, Sehulster LM, Dreesman GR, et al. Effect of alkaline glutaraldehyde on hepatitis B virus antigen. Eur J Clin Microbiol 1983;2:316–320.
193. Passagot J, Crance JM, Biziagos E, et al. Effect of glutaraldehyde on the antigenicity and infectivity of hepatitis A virus. J Virol Methods 1987;16:21–28.
194. Deva AK, Vickery K, Zou J, et al. Evaluation of an in-use testing method for evaluating disinfection of surgical instruments using the duck hepatitis B model. J Hosp Infect 1996;33:119–130.
195. Sattar AS, Raphael RA, Lochman H, et al. Rotavirus inactivation by chemical disinfectants and antiseptics used in hospitals. Can J Microbiol 1983;29:1464–1469.
196. Doultree JC, Druce JD, Birch CJ. Inactivation of feline calcivirus, a Norwalk surrogate virus. J Hosp Infect 1999;41:51–57.
197. Chan Myers H, Roberts C. Virucidal activity of o-phthalaldehyde solution against bovine viral diarrhea virus (BVDV). Paper presented at: The Association for Professionals in Infection Control and Epidemiology Conference; May 2002; Nashville, TN.
198. Roberts C, Chan Myers H. Virucidal activity of o-phthalaldehyde solution against duck hepatitis B virus. Paper presented at: The Association for Professionals in Infection Control and Epidemiology Conference; June 2001; Seattle, WA.
199. Linley E, Denyer S, McDonnell G. Use of hydrogen peroxide as a biocide: new consideration of its mechanism. J Antimicrob Chemother 2012;67:1589–1596.
200. Lever AM, Sutton SVW. Antimicrobial effects of hydrogen peroxide as an antiseptic and disinfectant. In: Ascenzi JM, ed. Handbook of disinfectants and antiseptics. New York: Marcel Dekker, 1996:159–176.
201. Block SS. Peroxygen compounds. In: Block SS, ed. Disinfection, sterilization and preservation. 5th ed. Philadelphia: Lippincott Williams & Wilkins, 2001:185–204.
202. Klopotek BB. Peracetic acid methods for preparation and properties. Chimica Oggi 1998;16:33–37.
203. Fridovich I. The biology of oxygen radicals. Science 1978;201:875–879.
204. Russell AD. Similarities and differences in the response of microorganism to biocides. J Antimicrob Chemother 2003;52:750–763.
205. Finnegan M, Linley L, Denyer SP, et al. Mode of action of H2O2 and other oxidizing agents: differences between liquid and gas forms. J Antimicrob Chemother 2010;65:2108–2115.
206. Hofmann J, Jusdt G, Pritzkow W, et al. Bleaching activators and mechanism of bleaching activation. J Prakt Chem 1992;334:293–297.
207. Malchesky PS. Medical applications of peracetic acid. In: Block SS, ed. Disinfection, sterilization and preservation. 5th ed. Philadelphia: Lippincott Williams & Wilkins, 2001:979–996.
208. Rutala WA, Gergen MF, Weber DJ. Sporicidal activity of chemical sterilants used in hospitals. Infect Control Hosp Epidemiol 1993;14:713–718
209. Holton J, McDonald V. Efficacy of selected disinfectants against Mycobacteria and Cryptosporidium. J Hosp Infect 1994;27:105–115.
210. Rutala W, Weber D. Sterilization, high level disinfection and environmental cleaning. Infect Dis Clin North Am 2011;25:45–76.
211. Rogez-Kreuz C, Yousfir R, Soufflet C, et al. Inactivation of animal and human prions by hydrogen peroxide gas plasma sterilization. Infect Control Hosp Epidemiol 2009;30:769–777.
212. Commager H, Judis J. Mechanism of action of phenolic disinfectants. VI. Effects on glucose and succinate metabolism of Escherichia coli. J Pharm Sci 1965;54:1436–1439.
213. Denyer SP. Mechanism of action of biocides. Int Biodeterior Biodegradation 1990;26:89–100.
214. Fredrick JJ, Corner TR, Gerhardt P. Antimicrobial actions of hexachlorophene: inhibition of respiration in Bacillus megaterium. Antimicrob Agents Chemother 1974;6:712–721.
215. Hugo WB, Bowen JG. Studies on the mode of action of 4-ethylphenol on Escherichia coli. Microbios 1973;8:189–197.
216. Hugo WB, Bloomfield SF. Studies on the mode of action of the phenolic antibacterial agent fentichlor against Staphylococcus aureus and Escherichia coli. III. The effect of fentichlor on the metabolic activities of Staphylococcus aureus and Escherichia coli. J Appl Bacteriol1971;34:579–591.
217. Bloomfield SF. The effect of the phenolic antibacterial agent fentichlor on energy coupling in Staphylococcus aureus. J Appl Bacteriol 1974;37:117–131.
218. Goddard PA, McCue KA. Phenolic compounds. In: Block SS, ed. Disinfection, sterilization and preservation. 5th ed. Philadelphia: Lippincott Williams & Wilkins, 2001:255–281.
219. Kroll RG, Anagnostopoulos GD. Potassium leakage as a lethality index of phenol and the effect of solute and water activity. J Appl Bacteriol 1981;50:139–147.
220. Gale EF, Taylor ES. Action of tyrocidin and some detergent substances in releasing amino acids from the internal environment of Streptococcus faecalis. J Gen Microbiol 1947;1:77–84.
221. Judis J. Studies on the mechanism of action of phenolic disinfectants. I. Release of radioactivity from 14C-labelled Escherichia coli. J Pharm Sci 1962;51:261–265.
222. Judis J. Studies on the mechanism of action of phenolic disinfectants. II. Patterns of release of radioactivity from Escherichia coli labeled by growth on various compounds. J Pharm Sci 1963;52:126–131.
223. Joswick HL, Corner TR, Silvernale JN, et al. Antimicrobial action of hexachlorophene: release of cytoplasmic materials. J Bacteriol 1971;108:492–500.
224. Hugo WB, Bloomfield SF. Studies on the mode of action of the phenolic antibacterial agent fentichlor against Staphylococcus aureus and Escherichia coli. II. The effect of fentichlor on the bacterial membrane and the cytoplasmic constituents of the cell. J Appl Bacteriol1971;34:569–578.
225. Rubin J. Mycobacteriocidal disinfection and control. In: Block SS, ed. Disinfection, sterilization, and preservation. 4th ed. Philadelphia: Lea & Febiger, 1991:331–384.
226. Russell AD, Chopra I. Sporostatic and sporicidal agents: their properties and mechanism of action. In: Understanding antibacterial action and resistance. 2nd ed. London: Ellis Horwood, 1996:150–171.
227. Bancroft WD, Richter GH. The chemistry of disinfection. J Phys Chem 1931;35:511–530.
228. Hugo WB. Disinfection mechanisms. In: Russell AD, Hugo WB, Ayliffe GA, eds. Principles and practices of disinfection, preservation and sterilization. 3rd ed. Oxford: Blackwell Scientific Publications, 1999:258–283.
229. Bach D, Lambert J. Action de quelques antiseptiques sur les dehydrogenase du staphylocoque dore. Compt Rend Soc Biol (Paris) 1937;126:298–300.
230. Bach D, Lambert J. Action de quelques antiseptiques sur les dehydrogenase du staphylocoque dore; activants le glucose, l’acide formique et un certain nombre d’autres substrates. Compt Rend Soc Biol (Paris) 1937;126:300–302.
231. Regos J, Hitz HR. Investigations on the mode of action of triclosan, a broad spectrum antimicrobial agent. Zentralbl Bakteriol 1974;226:390–401.
232. McMurray LA, Oethinger M, Levy SB. Triclosan targets lipid synthesis. Nature 1998;394:531–532.
233. McMurray LA, McDermott PT, Levy SB. Genetic evidence that inhA of Mycobacterium smegmatis is a target for triclosan. Antimicrob Agents Chemother 1999;43:711–713.
234. Slater-Radosti C, Van Aller G, Greenwood R. Biochemical and genetic characterization of the action of triclosan on Staphylococcus aureus. J Biol Chem 2001;48:1–6.
235. Heath RJ, Yu Y-T, Shapiro MA, et al. Broad spectrum antimicrobial biocides target the FABI component of fatty acid synthesis. J Biol Chem 1998;46:30316–30320.
236. Heath RJ, Ronald JR, Holland DR, et al. Mechanism of triclosan inhibition of bacterial fatty acid synthesis. J Biol Chem 1999;274:11110–11114.
237. Waller RF, Keeling PJ, Donald RG, et al. Nuclear-encoded proteins target the plasmid in Toxoplasma gondii and Plasmodium falciparum. Proc Natl Acad Sci USA 1998;95:12352–12357.
238. Perozzo R, Kuo M, Sidhu A. Structural elucidation of the specificity of the antibacterial agent triclosan for malarial enoyl acyl carrier protein reductase. J Biol Chem 2002; 277:13106–13114.
239. Bruch M. Chlorxylenol: an old-new chemical. In: Ascenzi JM, ed. Handbook of disinfectants and antiseptics. New York: Marcel Dekker, 1996:265–294.
240. Rutala WA, Clontz EP, Weber DI, et al. Disinfection practices for endoscopes and other semicritical items. Infect Control Hosp Epidemiol 1991;12:282–288.
241. Martin LS, et al. Disinfection and inactivation of human T-lymphotropic virus type III/lymphadenopathy-associated virus. J Infect Dis 1985;2:400–403.
242. Druce JD, Jardine D, Locarnini SA, et al. Syringe cleaning techniques and transmission of HIV. AIDS 1995;9:1105–1107
243. Regos J, Zak O, Solf R, et al. Antimicrobial spectrum of triclosan, a broad-spectrum antimicrobial agent for topical applications. II. Comparison with some other antimicrobial agents. Dermatologica 1979;158:72–79.
244. Schmid MB, Kaplan M. Reduced triclosan susceptibility in methicillin resistant Staphylococcus epidermidis. Antimicrob Agents Chemother 2004;48:1397–1399.
245. Suller MTE, Russell AD. Triclosan and antibiotic resistance in Staphylococcus aureus. J Antimicrob Chemother 2000;46:11–18.
246. Cookson BD, Farrelly H, Stapleton P, et al. Transferable resistance to triclosan in MRSA. Lancet 1991;337:1548–1549.
247. Webster J. Hand washing in a neonatal intensive care nursery: product acceptability and effectiveness of chlorhexidine gluconate and triclosan 1%. J Hosp Infect 1992;21:137–141.
248. Bhargava HN, Leonard PA. Triclosan: applications and safety. Am J Infect Control 1996;24:209–218.
249. Larson EL. Guidelines for the use of topical antimicrobial agents. Am J Infect Control 1988;8:253–266.
250. Sharma S, Ramya TNC, Surolia A, et al. Triclosan as a systemic antibacterial agent in a mouse model of acute bacterial challenge. Antimicrob Agents Chemother 2003;47:3859–3866.
251. Saunders KA, Greenman J, McKenzie C. Ecological effects of triclosan and triclosan monophosphate on defined mixed cultures of oral species grown in continuous culture. J Antimicrob Chemother 2000;48:447–452.
252. McBain AJ, Bartolo RG, Catrenich CF, et al. Effects of triclosan-containing rinse on the dynamics and antimicrobial susceptibility of in vitro plaque ecosystems. Antimicrob Agents Chemother 2003;11:3531–3538.
253. Simmons BP. Guideline for hospital environmental control. Am J Infect Control 1983;11:97–115.
254. Boyce JM, Pittet D. Guideline for hand hygiene in health care settings. MMWR Recomm Rep 2002;51:1–44.
255. Rutala WA, Cole EC. Antiseptics and disinfectants—safe and effective? Infect Control 1984;5:215–218.
256. Shickman MD, Guze LB, Pearce ML. Bacteremia following cardiac catheterization. N Engl J Med 1959;260:1164–1166.
257. Ehrenkranz NJ, Bolyard EA, Wiener M, et al. Antibiotic-sensitive Serratia marcescens infections complicating cardiopulmonary operations: contaminated disinfectant as a reservoir. Lancet 1980;2:1289–1292.
258. Merianos JJ. Surface-active agents. In: Block SS, ed. Disinfection, sterilization and preservation. 5th ed. Philadelphia: Lippincott Williams & Wilkins, 2001:283–320.
259. Tischer M, Pradel G, Ohlsen K, et al. Quaternary ammonium salts and their antimicrobial potential: targets or nonspecific interactions. Chem Med Chem 2012;7:22–31.
260. Rutala WA, Cole EC. Ineffectiveness of hospital disinfectants against bacteria: a collaborative study. Infect Control 1987;8:501–506.
261. Cole EC, Rutala WA, Samsa GP. Disinfectant testing using a modified use-dilution method: collaborative study. J Assoc Off Anal Chem 1988;71:1187–1194.
262. Zhao T, Sun G. Hydrophobicity and antimicrobiology and activity of quaternary ammonium compounds. J Applied Micro 2008;104:824–830.
263. Sasatsu M, Shibata Y, Noguchi N, et al. High-level resistance to ethidium bromide and antiseptics in Staphylococcus aureus. FEMS Microbiol Lett 1992;93:109–114.
264. Russell AD. Plasmid and bacterial resistance to biocides. J Appl Microbiol 1997;82:155–165.
265. Russell AD. The role of plasmids in bacterial resistance to antiseptics, disinfectants and preservatives. J Hosp Infect 1985;6:9–19.
266. Moken MC, McMurray LM, Levy SB. Selection of multiple-antibiotic-resistant (mar) mutants of Escherichia coli by using the disinfectant pine oil: roles of the mar and acr AB loci. Antimicrob Agents Chemother 1997;41:2770–2772.
267. Price CTD, Singh VK, Jayaswal RK, et al. Pine oil cleaner resistant Staphylococcus aureus: reduced susceptibility for vancomycin and oxicillin and involvement of SigB. Appl Env Microbiol 2002;68:5417–5421.
268. Rutala W. Elaine L. Larson Lectureship. Disinfection and sterilization: from benchtop to bedside. Association for Professionals in Infection Control and Epidemiology (APIC) Annual Education 2012 Conference. http://www.unc.edu/depts/spice/dis/LarsonLect12NP. Accessed November 15, 2012.
269. Sheldon AT. Antiseptic “resistance”: real or perceived threat? Clin Infect Dis 2005;40:1650–1656.
270. Levy SB. Antimicrobial household products: cause for concern. Emerg Infect Dis 2001;7:512–515.
271. Romanova N, Favrin S, Griffiths MW. Sensitivity of Listeria monocytogenes to sanitizers used in the meat processing industry. Appl Environ Microbiol 2002;68:6405–6409.
272. Tumah HN. Bacterial biocide resistance. J Chemother 2009;21:5–15.
273. Jalier V, Nikaido H. Mycobacterial cell wall: structure and role in natural resistance to antibiotics. FEMS Microbiol Lett 1994;123:11–18.
274. Romanova NA, Wolffs PF, Brovko LY, et al. Role of efflux pumps in adaptation and resistance of Listeria monocytogenes to benzalkonium chloride. Appl Environ Microbiol 2006;72:3498–3503.
275. Huet AA, Raygada JL, Mendiratta K, et al. Multidrug efflux pump overexpression in Staphylococcus aureus after single and multiple in vitro exposures to biocides and dyes. Microbiology 2008;154:3144–3153.
276. Russell AD. Mechanism of bacterial resistance to biocides. Int Biodeterior Biodegradation 1995;36:247–265.
277. To MS, Favrin S, Romanova N, et al. Post-adaptational resistance to benzalkonium chloride and subsequent physiochemical modifications of Listeria monocytogenes. Appl Env Microbiol 2002; 68:5258–5264.
278. Ogase HI, Nigai I, Kameda K, et al. Identification and quantitative analysis of degradation products of chlorhexidine and chlorhexidine-resistant bacteria with three-dimensional high performance liquid chromatography. J Appl Bacteriol 1992;73:71–78.
279. Costerton JW, Cheng KJ, Geesey GG, et al. Bacterial biofilms in nature and disease. Ann Rev Microbiol 1987;41:435–464.
280. Costerton JD, Lewandowski Z, DeBeer D, et al. Biofilms, the customized niche. J Bacteriol 1994;176:2137–2142.
281. Brown MRW, Gilbert P. Sensitivity of biofilms to antimicrobial agents. J Appl Bacteriol Symp Suppl 1993;74:87S–97S.
282. Nye K, Chadha DK, Hodgkin P, et al. Mycobacterium chelonei isolation from bronchoalveolar lavage fluid and its practical implications. J Hosp Infect 1990;16:257–261.
283. Fraser VJ, Jones M, Murray PR, et al. Contamination of flexible fiberoptic bronchoscopes with Mycobacterium chelonae linked to an automated bronchoscope disinfection machine. Am Rev Resp Dis 1992;145:853–855.
284. Griffiths PA, Babb JR, Bradley CR, et al. Glutaraldehyde-resistant Mycobacterium chelonae from endoscope washer disinfectors. J Appl Microbiol 1997;82:519–526.
285. Lynam P, Babb JR, Fraise AP. Comparison of the mycobacteriocidal activity of 2% alkaline glutaraldehyde and “Nu-Cidex” (0.35% peracetic acid). J Hosp Infect 1995;30:237–240.
286. Cabrera-Martinez R-M, Setlow B, Setlow P. Studies on the mechanism of the sporicidal action of ortho-phthalaldehyde. J Appl Microbiol 2002;92:675–680.
287. Vohra P, Poxton I. Efficacy of decontaminants and disinfectants against Clostridium difficile. J Med Microbiol 2011;60:1218–1224.
288. Powers EGM, Dancer BN, Russell AD. Emergence of resistance to glutaraldehyde in spores of Bacillus subtilis. FEMS Microbiol Lett 1988;50:223–226.
289. Knott AG, Russell AD, Dancer BN. Development of resistance to biocides during sporulation of Bacillus subtilis. J Appl Bacteriol 1995;79:492–498.
290. Bloomfield SF, Arthur M. Interaction of Bacillus subtilis spores with sodium hypochlorite, sodium dichloroisocynaurate. J Appl Bacteriol 1992;72:166–172.
291. Bloomfield SF, Arthur M. Mechanism of inactivation and resistance of spores to chemical biocides. J Appl Bacteriol Symp Suppl 1994;76:91S–104S.
292. Setlow B, Setlow P. Binding of small, acid-soluble spore proteins to DNA plays a significant role in the resistance of Bacillus subtilis spores to hydrogen peroxide. Appl Environ Microbiol 1993;59:3418–3423.
293. Sabli MZH, Setlow P, Waites WM. The effect of hypochlorite on spores of Bacillus subtilis lacking small acid-soluble proteins. Lett Appl Microbiol 1996;22:405–507.
294. Bay D, Turner R. Spectroscopic analysis of the intrinsic chromophores within small resistance protein SugE. Biochem Biophys Acta 2011;1808:2233–2244.
295. Walsh SE, Maillard JY, Russell AD. Ortho-phthalaldehyde: a possible alternative to glutaraldehyde for high level disinfection. J Appl Microbiol 1999;86:1039–1046.
296. Guerin-Mechin L, Dubois-Brissonnet F, Heyd B, et al. Quaternary ammonium compound stress induces specific variation in fatty acid composition of Pseudomonas aeruginosa. Int J Food Microbiol 2000;55:157–159.
297. Bridier A, Briandet R, Thomas V, et al. Comparative biocidal activity of paracetic acid, benzalkonium chloride and OPA on 77 bacterial strains. J Hosp Infect 2011;78:208–213.
298. Gilbert P, Brown MRW. Some perspectives on preservation and disinfection in the present day. Int Biodeterior Biodegradation 1995;36:219–226.
299. Kolawole DO. Resistance mechanism of mucoid-grown Staphylococcus aureus to the antibacterial action of some disinfectants and antiseptics. FEMS Microbiol Lett 1984;25:205–209.
300. Braoudaki M, Hilton AC. Adaptive resistance to biocides in Salmonella and Escherichia coli 0157 and cross-resistance to antimicrobial agents. J Clin Microbiol 2004;42:73–78.
301. Chuanchuen R, Narasaki CT, Schweizer HP. The MexJK efflux pump of Pseudomonas aeruginosa requires OprM for antibiotic efflux but not for efflux of triclosan. J Bacteriol 2002;184:5036–5044.
302. Langsrud S, Sundheim G, Borgman-Strahsen R. Intrinsic and acquired resistance to quaternary ammonium compounds in food related Pseudomonas spp. J Appl Microbiol 2003;95:874–882.
303. Heir E, Sundheim G, Holck AL. The Staphylococcus qacH gene product: a new member of the SMR family encoding multidrug resistance. FEMS Microbiol Lett 1998;163:49–56.
304. Heir E, Sundheim G, Holck AL. The qacG gene on plasmid pST94 confers resistance to quaternary ammonium compounds in staphylococci isolated from the food industry. J Appl Microbiol 1999;86:378–388.
305. Sidhu MS, Heir WE, Sørum H, et al. Genetic linkage between resistance to quaternary ammonium compounds and β-lactam antibiotics in food-related Staphylococcus spp. Microb Drug Resist 2001;7:363–371.
306. Sidhu MS, Heir E, Leegaard T, et al. Frequency of disinfectant resistance genes and genetic linkage with β-lactamase transposon Tn552 among clinical staphylococci. Antimicrob Agents Chemother 2002;46:2797–2803.
307. Leelaporn A, Firth N, Paulsen IT, et al. Multidrug resistance plasmid pSK108 from coagulase-negative staphylococci; relationship to Staphylococcus aureus qacC plasmids. Plasmid 1995;34:62–67.
308. Littlejohn TG, DiBerardino D, Messerotti LJ, et al. Structure and evolution of a family of genes encoding antiseptic and disinfectant resistance in Staphylococcus aureus. Gene 1991;101:59–66.
309. Paulsen IT, Brown MH, Littlejohn TG, et al. Multidrug resistance proteins QacA and QacB from Staphylococcus aureus: membrane topology and identification of residues involved in substance specificity. Proc Natl Acad Sci USA 1996;93:3630–3635.
310. Bjorland J, Steinum T, Sunde M, et al. Novel plasmid-borne qacJ mediates resistance to quaternary ammonium compounds in equine Staphylococcus aureus, Staphylococcus simulans, and Staphylococcus intermedius. Antimicrob Agents Chemother 2003;47:3046–3052.
311. Ciusa ML, Furi L, Knight D, et al. A novel resistance mechanism to triclosan that suggests horizontal gene transfer and demonstrates a potential selective pressure for reduced biocide susceptibility in clinical strains of S. aureus. Int J Antimicrob Agents 2012;40:210–220.
312. Lyon BR, Skurray RA. Antimicrobial resistance of Staphylococcus aureus: genetic basis. Microbiol Rev 1987;51:88–134.
313. Berq T, Firth N, Apisirdiej S, et al. Complete nucleotide sequence of pSK41: evolution of staphylococcal conjugative multiresistance plasmids. J Bacteriol 1998;180:4350–4359.
314. Alam MM, Kobayashi N, Uehara N, et al. Analysis of distribution and genomic diversity of high-level antiseptic resistance genes qacA and qacB in human clinical isolates of Staphylococcus aureus. Microb Drug Res 2003;9:109–121.
315. Lemaitre JP, Echchannaoui H, Michaut G, et al. Plasmid-mediated resistance to antimicrobial agents among listerae. J Food Prot 1998;61:1459–1464.
316. Huillet E, Larin S, Pardon P, et al. Identification of a new locus in Listeria monocytogenes involved in cellobiose dependent repression of hly expression. FEMS Microbiol Lett 1999;174:265–272.
317. Mereghetti L, Quentin R, Marquet-Van Der Mee N, et al. Low sensitivity of Listeria monocytogenes to quaternary ammonium compounds. Appl Env Microbiol 2000;66:5083–5086.
318. Rutala WA, Stergel MM, Sarubbi FA, et al. Susceptibility of antibiotic-susceptible and antibiotic-resistant hospital bacteria to disinfectants. Infect Control Hosp Epidemiol 1997;18:417–421.
319. Cole EC, Addison RM, Rubino JR, et al. Investigation of antibiotic and antibacterial agent cross-resistance in target bacteria from homes of antibacterial users and nonusers. J Appl Microbiol 2003;95:664–676.
320. Jones RD. Bacterial resistance and topical antimicrobial wash products. Am J Infect Control 1999;27:351–363.
321. Russell AD. Do biocides select for antibiotic resistance? J Pharm Pharmacol 2000;52:227–233.
322. Fanning S. Altered tolerance to biocides: links to antibiotic resistance? Paper presented at: International Association of Food Protection (IAFP), European Symposium on Food Safety; 2011; The Netherlands. http://www.foodprotection.org/events/european-symposia/11Ede/Fanning.pdf. Accessed November 15, 2012.
323. Alexander L. Nosocomial infections. http://www.netcegroups.com/372/course_9447.pdf. Accessed November 15, 2012.
324. Andersen B, Banrud H, Boe E, et al. Comparison of UV C light and chemicals for disinfection of surfaces in hospital isolation units. Infect Control Hosp Epidemiol 2006;27:729–734.
325. Rutala W, Weber D. Guideline for disinfection and sterilization of prion contaminated medical instruments. SHEA guidelines. Infect Control Hosp Epidemiol 2010;31:107–117.
326. The Joint Commission. Joint Commission 2013 National Patient Safety Goals. January 2013. http://www.jointcommission.org/hap_2013_npsg/. Accessed November 15, 2012.