Toshiki Mizobe
Daniel I. Sessler
History
Well before the dawn of clinical thermometry, the temperature course of diseases such as malaria and enteric fever (typhoid and brucellosis) were well described in the Corpus Hippocraticum (circa 370–460 BC) (1). However, it wasn't until the 1590s when the astronomer Galileo invented the thermometer (or thermoscope) that humans could record temperature. His thermoscope was an open thermometer subject to atmospheric pressure. In 1612, Sanctorius of Justipolitanus at Padua first applied the thermometer to the human body. A major advance in temperature monitoring arrived in 1714 when Gabriel Fahrenheit invented the mercury thermometer (2). His scale, the Fahrenheit scale, was and remains commonly used in England and the United States. However, another scale, the Centigrade scale, invented by Anders Celsius in 1742, became popular in France and Germany. On the original Celsius scale, 0° was the temperature at which water boiled, while 100° was the temperature at which ice melted. Linnaeus reversed the scale in 1750.
Carl Reinhold Wunderlich developed the thermometer that was used clinically in medicine for over 130 years (3). It differed from previous ones in preserving the maximum temperature in any given session that, presumably, best represents core temperature. He published Das Verhalten der Eigenwarme in Krankheiten in 1868, in which he suggested that fever is not a disease, but rather a sign of disease. His thermometers were 22.5 cm long and took 20 minutes to register temperature. Measurements were made in the axilla with four to six daily observations made on patients with fever. After obtaining several million observations on approximately 25,000 patients, he established the fever patterns of diseases such as typhoid fever, various internal diseases, and pregnancy. He also established the average normal body temperature as 98.6°F (37°C). He concluded, “A physician who practiced medicine without employing the thermometer was like a blind man endeavoring to distinguish colors by feeling.”
Wunderlich's findings were supported enthusiastically in Europe and in the United States. It was already recognized that thermometer use facilitated the recognition of diseases and allowed physicians to understand their natural progression. However, the curved and nonregistering thermometers then in use were so cumbersome that more portable instruments were required. With the size reduced by Wunderlich, clinical thermometry became a routine part of medical practice. By the 1860s, the axilla had become universally adopted as a convenient and reliable point of measurement. Once the use of alcohol and other germicidal agents became common in the late 1890s, sublingual placement replaced the axilla as the most popular measurement site.
In 1875 von Liebermeister first hypothesized that body temperature is regulated similarly in both healthy and ill people, but that fever occurs in illness because the body's internal thermostat is set to a higher temperature. Edward Seguin and William Draper introduced thermometry and patient charting in New York City hospitals in the mid-1860s. Medical thermometry quickly spread from hospitals to general practitioners, and from there to nurses and family. Mothers who could read a thermometer rendered invaluable service to their families in times of illness and provided important information for physicians. Seguin even concluded that thermometry is not only knowledge, but also social power.
Types of Thermometers
Mercury-in-glass Maximum
This familiar design consists of a mercury-filled bulb connected to a thin glass tube. As the body warms the mercury in the bulb, thermal expansion causes the mercury to move up the tube. After maximal thermal equilibrium is reached (which may require 4 minutes in the mouth and 10 minutes in the axilla), bulb temperature equals the body temperature. A constriction near the bulb holds the position of mercury when the thermometer is removed from the body, thus preserving the maximum temperature reading. After the thermometer is shaken to return mercury to the bulb, another measurement can be performed.
The advantages of mercury-in-glass thermometers are that they do not require electric power, accuracy is easily confirmed by calibration, and environmental conditions do not affect clinical measurements (4). The disadvantages of these thermometers include that they require a long equilibration time, they cannot be used orally except in cooperating adults, and they present a risk of mercury and glass exposure if damaged during storage and clinical use. For instance, about 1,600 glass thermometers were reported broken at a Glasgow Children's Hospital during a 6-month period. Spilled mercury disperses into tiny droplets that are difficult to see and easily become entrapped and emit a toxic vapor that can be inhaled or absorbed through the skin; the vapor may persist for months or even years (5).
Electronic Contact
A thermistor is an electrical resistor in which resistance changes rapidly in response to temperature fluctuation. In general, resistance decreases exponentially with increasing temperature. Electrical power is required for measuring the resistance and calibrating stored data in order to convert resistance data into temperature data. Thermocouples measure temperature by evaluating the tiny electrical potential (voltage) produced by the junction of two metals. Both thermistors and thermocouples are inexpensive, with a manufacturing cost as low as a few cents each, and are highly accurate. These thermometers are, by far, the most commonly used at present.
Most products display the maximum temperature and provide an audible tone when the temperature increase slows to 0.1°C in a preset time, generally in 8 seconds. Underreading occurs occasionally, especially when measuring axillary temperature where the dry skin requires a longer time to achieve thermal equilibrium with the sensor. Some devices provide a predictive mode that tracks the changing resistance to estimate the final temperature. This results in a rapid temperature reading within several seconds; however, accuracy is worse in the predictive mode than in the monitoring mode.
Infrared Aural Canal (“Tympanic”)
Optical infrared emissions sensors can be used to measure surface temperature. Industrially, these devices are used to measure the temperature of objects that would be hard to otherwise measure, such as the temperature of molten steel. However, they are also used medically to measure skin temperature. The magnitude and spectrum of the infrared energy emitted depends on the local temperature, efficiency of the surface for radiating electromagnetic radiation (emissivity), filtering effect of any optical components, and sensor temperature. The resulting signal is converted into a temperature.
International standards permit a lower accuracy (±0.2°C) in laboratory testing of clinical infrared thermometers than the corresponding standards of conventional contact thermometers (±0.1°C). Even larger errors are permitted when the ambient temperature is out of normal room temperature, about 18°C to 26°C. Although rapid response is a major advantage of this type of device, users must ensure that the aural canal is not obscured by cerumen and that the ear canal is straightened by manipulating the ear lobe so that the sensor can point directly at deeper sections of the canal, which are nearer to the core temperature (6). Even though infrared aural canal thermometers are often labeled as “tympanic,” they do not actually “see” the tympanic membrane but extrapolate tympanic or core temperature from the skin temperature in the aural canal.
Sites of Temperature Measurement
The core thermal compartment is composed of well-perfused tissues with a temperature uniform and high compared with the rest of the body. Core temperature is considered the average temperature of the core compartment; it is not the highest temperature in the body, which is probably in the brain or liver. Core temperature can be evaluated in the pulmonary artery, distal esophagus, tympanic membrane, or nasopharynx. Even during rapid thermal perturbations (e.g., cardiopulmonary bypass), these temperature-monitoring sites remain reliable. Core temperature can be estimated with reasonable accuracy using oral, axillary, rectal, and bladder temperatures, except during extreme thermal perturbations (7,8).
1. Oral: The most common site of temperature measurement is probably under the tongue. Unfortunately, hot and cold drinks, open-mouth versus closed-mouth breathing, and tachypnea all affect oral temperature readings. However, nasogastric tubes do not interfere with oral temperature measurement (9,10,11). Carefully performed oral temperatures are reasonably accurate.
2. Axillary: Axillary temperature measurement is suited for intermittent monitoring, though the accuracy is worse than at other sites, probably due to probe positioning (7). Axillary measurements are, nonetheless, usually suitable if care is taken to position the probe over the axillary artery while keeping the arm at the patient's side.
3. Rectal: Rectal temperature measurement is most accurate when the sensor is placed more than 10 cm into the rectum. Rectal temperature correlates well with distal esophageal, bladder, and tympanic temperature, although it typically slightly exceeds core value. Also, it is the slowest to respond to rapid changes in body temperature (12). Consequently, rectal readings are often erroneous—sometimes by many degrees—during heat stroke, malignant hyperthermia, and other situations where core temperature changes rapidly. It is thus a poor choice when a purpose of monitoring is to detect rapid perturbations.
4. Tympanic: Actual tympanic membrane measurements, obtained with a thermocouple or thermistor, are highly accurate core temperatures (13). During induced hypothermia, tympanic temperature is much closer to brain temperature than other measurement sites (14).
5. Infrared aural canal: Infrared aural canal temperatures do not actually evaluate the tympanic membrane and are far less accurate. These measurements may be adequate when patients have wide ear canals (large circumference of the ear canal) and good visibility deep in the canal. However, values deviate considerably when hair or cerumen even partially occludes the canal—as is typical. Furthermore, fanning the face has been reported to lower the tympanic temperature. In general, infrared aural canal “tympanic” thermometers are insufficiently accurate for routine clinical measurements.
6. Distal esophageal: Distal esophageal temperature is an accurate monitoring site for core temperature when patients are anesthetized or sedated, particularly because this site is resistant to artifact (15). The most precise measurement is performed when the probe is 45 cm from the nose in adults. The temperature measurements taken in the proximal and midesophagus are influenced by the ambient air because they are near the trachea and bronchi. Transesophageal echocardiography (TEE) may affect the esophageal temperature because of the heat emitted by the probe.
7. Nasopharyngeal: Nasopharyngeal temperature monitoring is often performed during general anesthesia. The drawbacks of this site are that the measurements are affected by the probe position and that nasopharyngeal bleeding might occur. Nonetheless, it is an excellent site and usually provides accurate and precise measurements of core temperature.
|
|
|
Figure 24.1. Control of autonomic and behavioral thermoregulatory defenses. Efferent defenses can be broadly divided into autonomic responses (e.g., sweating and shivering) and behavioral responses (e.g., closing a window, putting on a sweater). Autonomic responses depend largely on core temperature and are mediated largely by the anterior hypothalamus. In contrast, behavioral responses are mostly determined by skin temperature and are controlled by the posterior hypothalamus. |
8. Urinary bladder: Urinary bladder temperature is measured by placing a probe in a Foley catheter. The measurement is affected by urinary flow during cardiopulmonary bypass and does not provide an accurate temperature measurement when urine flow is low (16). Under other circumstances, though, it is a reasonably accurate site.
9. Pulmonary artery: Pulmonary artery temperature is measured by a sensor located at the distal end of the balloon-tipped, flow-directed pulmonary artery catheter. It is generally considered the single, best core temperature measurement site, although obviously available in only a tiny fraction of patients. It is worth noting that brain temperature is slightly greater than pulmonary artery temperature, because core temperature at the pulmonary artery is the average value of the deep body structures. The gradient between core and brain temperatures tends to increase with fever and active cooling.
Thermoregulation
Precise control of core temperature is maintained by a powerful thermoregulatory system incorporating afferent inputs, central control, and efferent defenses. Efferent defenses can be broadly divided into autonomic responses (e.g., sweating and shivering) and behavioral responses (e.g., closing a window, putting on a sweater). Autonomic responses depend largely on core temperature and are mostly mediated by the anterior hypothalamus. In contrast, behavioral responses are roughly 50% determined by skin temperature and are controlled by the posterior hypothalamus (Fig. 24.1) (17,18).
|
|
|
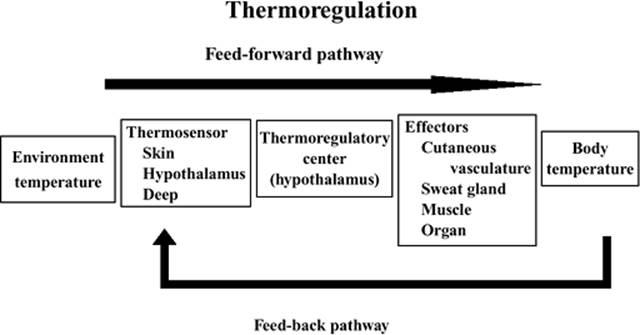
Figure 24.2. Signals from the cutaneous thermosensors detecting changes in environmental temperature are transmitted to the thermoregulatory center. The effectors that receive efferent signals control thermoregulatory homeostasis (feed-forward). Also, the deep thermosensors that detect changes in body temperature transmit the afferent signals to the central controller (feed-back). |
Thermoregulation is maintained by feed-forward and feed-back pathways. The signal from the cutaneous thermosensors detecting the change in environmental temperature is transmitted to the thermoregulatory center in the hypothalamus. The effectors receiving efferent signals from the thermoregulatory center control thermoregulatory homeostasis. Also, the deep thermosensors detecting the change in body temperature transmit the afferent signals to the center. Recent studies suggest that the feed-forward pathway may be more important than previously appreciated (Fig. 24.2).
Afferent Input
Dual detectors—cold sensors with myelinated A-delta fibers and warm sensors with unmyelinated C-fibers—exist in the subcutaneous layer of the skin. The peripheral temperature sensors are free nerve endings with no specialized structure. They differ in the temperature range over which they operate and in their responses to temperature change. The response curve of thermosensors for a constant temperature exhibits a maximal discharge frequency in the range of 25°C to 30°C for cold sensors and around 40°C for warm sensors (static response). Cold sensors react to cooling with a transient increase in activity (dynamic response) and are inhibited during warming (Fig. 24.3). Warm sensors demonstrate dynamic activation during a temperature increase and are inhibited during cooling.
The distribution density of cold sensors is several times greater than warm sensors in the skin. These peripheral sensors detect not only the body temperature, but also the environmental temperature. Most thermal input is conducted along the spinothalamic tracts, although both afferent and efferent thermal signals are diffusely distributed within the neuraxis. Interestingly, signals from nociceptive receptors are conducted along almost the same part of the spinothalamic tracts (19).
Thermosensors are also located in brain areas such as the hypothalamus, midbrain, and spinal cord, and in deep thoracic and abdominal tissues. These deep sensors detect core body temperature. The dynamic responses shown by peripheral thermosensors are not observed in the thermosensors in the brain. It is likely that there are also thermoreceptors in the thorax and abdomen, but their specific contributions in humans remain unknown.
|
|
|
Figure 24.3. The response curve of thermosensors for a constant temperature exhibits a maximal discharge frequency in the range of 25°C to 30°C for cold sensors and around 40°C for warm sensors (static response). Cold sensors react to cooling with a transient increase in activity (dynamic response) and are inhibited during warming. Warm sensors demonstrate dynamic activation during a temperature increase and are inhibited during cooling (dynamic response). Change is seen in the impulse frequency while temperature turns from T1 to T2 and back to T1 again. (Modified from Hensel H. Thermoreception and Temperature Regulation. London: Academic Press; 1981.) |
Central Control
Thermal afferent signals are integrated at numerous levels within the neuraxis, including the spinal cord and brainstem. The hypothalamus is undoubtedly the dominant and most precise controller of body temperature, although the spinal cord dominates in birds. As there are few cold sensors in the hypothalamus, warm sensors are thought to send the stimulatory and inhibitory signals to the effectors.
Core temperature varies with a daily circadian rhythm (20). Normal temperature is altered slightly by factors such as age (0.5°C lower in the elderly), time of day (0.5°C higher in the afternoon), time of menstrual cycle in women (higher near ovulation), and exercise (21). Nonetheless, core temperature is normally controlled within a few tenths of a degree centigrade, virtually irrespective of the environment (22). Such precise control is maintained by a powerful thermoregulatory system incorporating afferent inputs, central control, and efferent defenses. The thresholds triggering thermoregulatory defenses are uniformly ~0.3°C greater during the follicular phase in women than in men (22), and are an additional ~0.5°C greater during the luteal phase (23). However, men and women regulate core body temperature with comparable precision, usually maintaining core temperature within a few tenths of a degree centigrade of the target temperature (Fig. 24.4).
|
|
|
Figure 24.4. The thresholds (triggering core temperatures) for the three major autonomic thermoregulatory defenses: Sweating, vasoconstriction, and shivering. Temperatures between the sweating and vasoconstriction threshold define the interthreshold range (temperatures not triggering autonomic responses). The thresholds are uniformly about 0.3°C greater during the follicular phase in women than in men, and are an additional ≈0.5°C greater during the luteal phase. However, men and women regulate core body temperature with comparable precision. Results are presented as means ± standard deviation. (From Lopez M, Sessler DI, Walter K, et al. Rate and gender dependence of the sweating, vasoconstriction, and shivering thresholds in humans. Anesthesiology. 1994;80:780–788, with permission.) |
Nonthermoregulatory cutaneous circulatory reflexes, including cardiopulmonary and arterial baroreceptor reflexes, modulate thermoregulatory vascular tone. The cardiopulmonary baroreceptor reflex controls peripheral vascular resistance in response to the change in blood volume through the central sympathetic nervous system and the renin–angiotensin pathway. The baroreceptor reflex is modified by the right atrial transmural pressure, which is the difference between central venous pressure and the intrathoracic pressure. Baroreflex loading by increased right atrial pressure in patients placed in the leg-up position results in an exaggeration of anesthesia-induced hypothermia because of attenuated peripheral vasoconstriction. In contrast, positive end-expiratory pressure (PEEP) ventilation decreases right atrial transmural pressure (RATP) and, as a consequence, attenuates perioperative hypothermia (Fig. 24.5) (24).
Efferent Responses
The balance between heat production and heat loss determines body temperature. Nonevaporative heat loss depends on the difference between skin temperature and environmental temperature. Given that skin blood flow is a primary determinant of skin temperature (e.g., increased blood flow results in increased skin temperature), skin vascular tonus can modulate heat loss. However, when environmental temperature exceeds body temperature, evaporative heat loss (sweating) is the only way to lose heat. Also, when environmental temperature is very low, heat production must increase if thermal homeostasis is to be maintained because maximal cutaneous vasoconstriction cannot compensate for the amount of heat loss. The thermoneutral zone can be defined as the range of environmental temperature over which cutaneous vasomotion alone can maintain body temperature (Fig. 24.6). The environmental temperature at which heat production increases—the lower critical temperature—is 29°C, while the temperature at which evaporative heat loss starts—the upper critical temperature—is 31°C.
|
|
|
Figure 24.5. The esophageal temperature (Tes) after induction of anesthesia. Positive end-expiratory pressure (PEEP: 10 cm H2O, P) or the leg-up position (L) was applied 10 minutes after induction of anesthesia. Baroreflex loading due to leg-up position exaggerates anesthesia-induced hypothermia because peripheral vasoconstriction is attenuated. Meanwhile, PEEP attenuated perioperative hypothermia because of stimulated vasoconstriction through baroreceptor unloading. Values are shown as mean ± standard of error. A significant difference compared with the control group (C). (From Nakajima Y, Mizobe T, Takamata A, et al. Baroreflex modulation of peripheral vasoconstriction during progressive hypothermia in anesthetized humans. Am J Physiol Regul Integr Comp Physiol. 2000;279:R1430–1436, with permission.) |
Sweating is mediated by postganglionic cholinergic nerves that terminate on sweat follicles (25). These follicles apparently have no purpose other than thermoregulation. In this regard, they differ from most other thermoregulatory effectors that appear to have been co-opted by the thermoregulatory system but continue to play important roles, for example, vasomotion in blood pressure control or skeletal muscles in postural maintenance. Heat exposure can increase cutaneous water loss from trivial amounts to 500 mL per hour. Sweat loss in trained athletes can even exceed 1 L/hour. In a dry, convective environment, sweating can dissipate enormous amounts of heat—perhaps up to ten times the basal metabolic rate.
|
|
|
Figure 24.6. The thermoneutral zone is defined as the environmental temperature zone in which skin vascular tonus alone can maintain body temperature. The environmental temperature at which heat production increases (the lower critical temperature) is 29°C, while the temperature at which evaporative heat loss starts (the upper critical temperature) is 31°C. |
As stated previously, sweating is the only mechanism by which the body can dissipate heat when environmental temperature exceeds core temperature. Fortunately, the process is remarkably effective, dissipating 0.58 kcal/g of evaporated sweat. Active precapillary vasodilation is mediated by a yet to be identified factor released from sweat glands and, thus, occurs synchronously with sweating. Recent evidence supports nitric oxide as the mediator of this response, although this theory remains controversial (26,27). Active dilation can increase cutaneous capillary flow enormously, perhaps to as much as 7.5 L/min (28). The purpose of this dilation, presumably, is to transport heat from muscles and the core to the skin surface where it can be dissipated to the environment by evaporation of sweat.
Cutaneous vasoconstriction is the first autonomic response to cold. Metabolic heat is lost primarily via convection and radiation from the skin surface, and vasoconstriction reduces this loss. Active arteriovenous shunt vasoconstriction is adrenergically mediated. The shunts are vessels 100 µm in diameter and convey 10,000 times as much blood as a comparable length of 10-µm capillaries (29). Anatomically, they are restricted to the fingers, toes, nose, and nipples. Despite this restriction, shunt vasoconstriction is among the most commonly used and important thermoregulatory defenses. The reason is that the blood traversing via shunts in the extremities must flow through the arms and legs, thus altering the heat content of these relatively large tissue masses. Local α-adrenergic sympathetic nerves mediate constriction in the thermoregulatory arteriovenous shunts, while circulating catecholamines only minimally affect flow. Roughly 10% of cardiac output traverses arteriovenous shunts; consequently, shunt vasoconstriction increases mean arterial pressure by approximately 15 mm Hg.
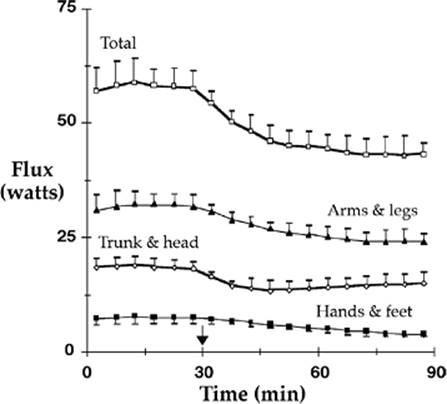
Thermoregulatory vasoconstriction reduces cutaneous heat loss. Although there is a major regional decrease in the distal extremities, these areas constitute a relatively small fraction of the body surface area. Nonetheless, heat loss does decrease throughout the extremities—including the proximal segments—because the entire limb cools when shunt flow is obliterated. In contrast, heat loss from the trunk decreases only slightly (Fig. 24.7). The overall reduction in heat loss that results from thermoregulatory vasoconstriction approximates 25%, which is similar to that provided by a single layer of most any insulator (30,31).
A major purpose of thermoregulatory vasoconstriction is to isolate core tissues from the environment, and thus restrict peripheral-to-core heat transfer. Normally, this is beneficial, but when patients are cooled therapeutically, thermoregulatory vasoconstriction slows transfer of heat from the skin surface to the core by trapping heat in peripheral tissues (32,33). The normal threshold (triggering core temperature) for vasoconstriction is approximately 36.5°C (22).
Thermoregulatory vasoconstriction affects blood pressure and heart rate. Vasoconstriction increases mean arterial pressure in awake subjects, and similar increases (14 ± 5 mm Hg) were observed in volunteers under various concentrations of isoflurane and desflurane with no dose dependency. Thus, the hypertensive response to thermoregulatory vasoconstriction remains well preserved, even during volatile anesthesia. In anesthetized subjects, the bradycardia to cold-induced hypertension was attenuated by the anesthesia, though awake subjects had a significant decrease in heart rate (Fig. 24.8) (33).
In humans, shivering is the final autonomic response that is activated to cold, and is generally only observed when behavioral responses and vasoconstriction fail to maintain an adequate core temperature. Shivering is an involuntary, thermogenic tonic tremor (34). Typically, vigorous shivering doubles the metabolic rate (35,36), although greater increases can be sustained briefly. The shivering threshold is approximately 35.5°C (22), which is about 1°C less than the vasoconstriction threshold. The low shivering threshold suggests that this response is activated only under critical conditions, rather than being a preferred means of maintaining core temperature. Shivering begins with the pectoralis muscles, but most shivering thermogenesis occurs in the extremities where the largest muscles are located. The muscular activity of shivering does not perform any work, so all expended energy is converted into heat. However, the ability of shivering thermogenesis to protect core temperature is limited by two factors. The first is that active muscle must be supplied with oxygen and metabolic substrates, both of which are conveyed by blood. This need for blood flow in distal muscles counteracts the efforts of vasoconstriction to constrain metabolic heat to the core thermal compartment, and, thus, establishes a large core-to-peripheral tissue-temperature gradient. The second limitation of shivering is that most heat is generated peripherally, and thus easily dissipates to the environment.
|
|
|
Figure 24.7. Cutaneous heat flux (heat loss) from the head and trunk (head, back, chest, and abdomen), legs and arms (upper arm, lower arm, thigh, and calf), and hands and feet. Total heat flux and flux from the arms and legs decreased about 25% after vasoconstriction was triggered by infusion of cold fluid at 30 elapsed minutes (arrow). Heat loss from the trunk and head decreased only 17%. In contrast, heat loss from the hands and feet decreased about 50%. Heat losses indicated in the lower three curves (arms and legs, trunk and head, hands and feet) comprise the total loss indicated at the top of the figure. Results are presented as means ± standard deviations. (From Sessler DI, Moayeri A, Støen R, et al. Thermoregulatory vasoconstriction decreases cutaneous heat loss. Anesthesiology. 1990;73:656–660.) |
|
|
|
Figure 24.8. Systolic arterial pressure (SAP), diastolic arterial pressure (DAP), mean arterial pressure (MAP), and heart rate (HR) before and after vasoconstriction. Each symbol represents one group of study subjects without anesthesia or with various concentrations of isoflurane or desflurane. Data are represented as means of the individual studies, with the average and standard deviations for the entire study population also shown. Asterisks (*) identify statistically significant differences from vasodilatation (P <0.001). (From Greif R, Laciny S, Rajek A, et al. Blood pressure response to thermoregulatory vasoconstriction during isoflurane and desflurane anesthesia. Acta Anaesthesiol Scand. 2003;47:847–852, with permission.) |
An obvious consequence of shivering is increased metabolic rate. This increase, which is analogous to exercise, provokes a substantial adrenergic response. For example, a reduction in core temperature of only 0.7°C increases norepinephrine concentration by 400% and oxygen consumption by 30%. When core temperature decreases 1.3°C, norepinephrine concentration increases 700% and oxygen consumption doubles. As might be expected, shivering is associated with peripheral vasoconstriction and hypertension—although heart rate remains unchanged, as do plasma epinephrine and cortisol concentrations (37).
In general, energy-efficient effectors such as vasoconstriction are maximized before metabolically costly responses such as shivering are initiated. The interaction between thermal input, central control, and effector responses is shown in Figure 24.9; this figure also shows the normal values for the major autonomic response thresholds.
In the elderly, vasoconstriction in response to cold exposure is reduced (38), and it is likely that this is a clinically important observation because vasoconstriction is the primary autonomic response to cold exposure. Similarly, the shivering threshold is significantly reduced in the elderly (39). Interestingly, abnormally reduced thresholds were not apparent in subjects less than 80 years of age, and even then only occurred in a fraction of the population (Fig. 24.10). These data suggest that age-related thermoregulatory impairment may not be common in people younger than 80 years. The data further suggest that impairment is not a linear function of age, but instead occurs unpredictably in a fraction of the elderly population.
|
|
|
Figure 24.9. A schematic illustrating thermoregulatory control mechanisms. Mean body temperature is the integrated thermal input from a variety of tissues including the brain, skin surface, spinal cord, and deep core structures. These are shown entering the hypothalamus (large square) from the left. However, thresholds usually are expressed in terms of core temperature. A core temperature below the thresholds for response to cold provokes vasoconstriction, nonshivering thermogenesis, and shivering. Core temperature exceeding the hyperthermic thresholds produces active vasodilation and sweating. No thermoregulatory responses are initiated when core temperature is between these thresholds; these temperatures identify the interthreshold range. The interthreshold range in humans is usually only about 0.2°C. (Threshold data from Lopez M, Sessler DI, Walter K, et al. Rate and gender dependence of the sweating, vasoconstriction, and shivering thresholds in humans. Anesthesiology. 1994;80:780–788. Figure from Sessler DI. Perioperative hypothermia. N Engl J Med. 1997;336:1730–1737, with permission.) |
|
|
|
Figure 24.10. The effect of aging on the shivering threshold. Fifteen patients less than 80 years old (58 ± 10 years) (mean ± standard deviation) shivered at 36.1 ± 0.6°C; in contrast, ten patients aged 80 years or older (89 ± 7 years) shivered at a significantly lower mean temperature, 35.2 ± 0.8°C (P <0.001). The shivering thresholds in seven of the ten patients 80 years or older was less than 35.5°C, whereas the threshold equaled or exceeded this value in all the younger patients. (From Vassilieff N, Rosencher N, Sessler DI, et al. The shivering threshold during spinal anesthesia is reduced in the elderly. Anesthesiology. 1995;83:1162–1166, with permission.) |
The most powerful inhibitors of thermoregulation are general anesthetics. These drugs have been used to facilitate the induction of hypothermia for cardiac surgery and during neurosurgery. In sufficient doses, especially when combined with neuromuscular blockers, anesthetics can essentially obliterate defenses against hypothermia. Volatile anesthetics, such as desflurane and sevoflurane, have relatively little effect on sweating: Even full anesthetic doses increase the sweating threshold by only about 0.5°C. In contrast, anesthetic gases markedly reduce the vasoconstriction and shivering thresholds. Interestingly, the threshold for each major cold defense is reduced synchronously, as if they are similarly controlled (Fig. 24.11).
Neuromuscular blockers are not believed to have any effect on central control of thermoregulatory responses. However, they obviously cause paralysis and thus prevent shivering. It is equally obvious that muscle relaxants can only be given in the context of general anesthesia or substantial sedation, and that paralyzed patients require mechanical ventilation. Muscle relaxants are a suitable treatment for shivering in the intensive care unit (40), but are not a substitute for adequate sedation and appropriate thermal management.
|
|
|
Figure 24.11. Concentration-dependent thermoregulatory inhibition by desflurane and isoflurane (halogenated volatile anesthetics), propofol (an intravenous anesthetic), and alfentanil (a µ-agonist opioid). General anesthetics are the most powerful known inhibitors of thermoregulatory control. The horizontal axis, in each case, spans a clinically relevant concentration range. The sweating (triangles), vasoconstriction (circles), and shivering (squares) thresholds are expressed in terms of core temperature at a designated mean skin temperature of 34°C. Anesthesia linearly, but slightly, increases the sweating threshold. In contrast, anesthesia produces substantial and comparable linear or nonlinear decreases in vasoconstriction and shivering thresholds. Typical anesthetic concentrations thus increase the interthreshold range (difference between the sweating and vasoconstriction thresholds) approximately 20-fold from its normal value near 0.2°C. Patients do not activate autonomic thermoregulatory defenses unless body temperature exceeds the interthreshold range; surgical patients are thus poikilothermic over a 3°C to 5°C range of core temperatures. Isoflurane 1% and desflurane 6% have comparable anesthetic potency. Error bars smaller than the data markers have been deleted. (From Sessler DI. Perioperative hypothermia. N Engl J Med. 1997;336:1730–1737, with permission.) |
Therapeutic Hypothermia
Systemic Consequences of Hypothermia
Patients undergoing mild hypothermia exhibit decreased heart rate, increased systemic vascular resistance, and normal stroke volume; consequently, mean arterial blood pressure in these patients remains normal (41). The electrocardiogram of hypothermic patients often displays a notch on the downstroke of the QRS complex (the Osborn wave) when core temperature reaches 33°C (42). However, cardiac arrhythmias are rarely seen during therapeutic hypothermia, even in patients with myocardial ischemia (43).
Overall, hypothermia has little effect on the respiratory system. Minute ventilation decreases during therapeutic hypothermia, but PaCO2 remains normal since the metabolic rate decreases as well (44). Although pneumonia is listed as one of the major risk factors of hypothermia, it is relatively uncommon in adult patients subjected to short-term (12–24 hours) therapeutic hypothermia (43).
The number of leukocytes decreases and becomes less effective during hypothermia, a situation that could increase the incidence of infection, especially pneumonia (45). Prolonged hypothermia also decreases the number and function of platelets and delays clotting time, thus increasing the risk of hemorrhage (46).
During induction of therapeutic hypothermia, potassium shifts from the plasma into cells, resulting in hypokalemia. However, caution should be used in correcting this condition because during rewarming, the shift of potassium back into the plasma might lead to life-threatening hyperkalemia. Phosphate concentration decreases similarly (47,48). Urine output increases at induction of hypothermia as a result of decreased reabsorption in the ascending loop of Henle. It normalizes, however, when the patient reaches the target temperature (49). Volume status, potassium, and phosphate concentrations thus each require careful monitoring during therapeutic hypothermia.
Gas solubility in blood increases as body temperature decreases. It is controversial whether blood gas measurements should be corrected for body temperature during therapeutic hypothermia. Keeping a constant pH, which is known as pH-stat management, decreases the cerebral infarction area in a rat model (44). Nevertheless, the clinical evidence during cardiopulmonary bypass favors α-stat management, in which temperature correction is not performed for blood gas measurement (50).
Gut motility decreases during hypothermia, which delays enteral feeding (45). In addition, blood glucose concentrations increase in response to reduced insulin secretion. Insulin administration is recommended to correct hypothermia-induced hyperglycemia, and should probably be titrated to maintain a blood glucose concentration between 80 and 110 mg/dL (51,52).
Pathology of Cerebral Ischemia and Mechanism of Therapeutic Hypothermia
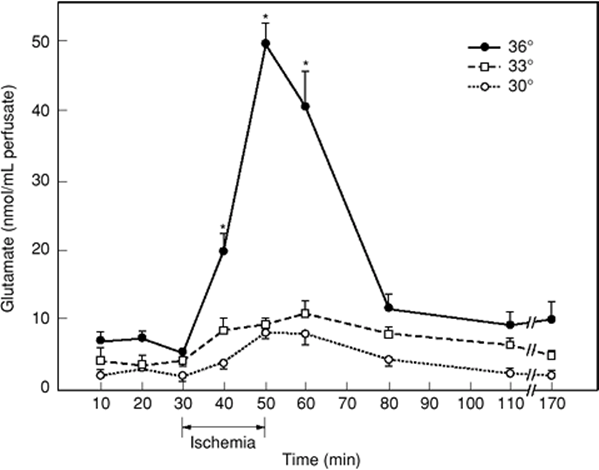
Cerebral injury occurs if inadequate blood flow to the brain occurs for more than 5 minutes (53). Cerebral ischemia produces a “whole-system failure” including adenosine triphosphate (ATP) energy depletion, ion pump failure, release of free radicals and excitotoxic neurotransmitters, and modulators such as glutamate and calcium (54,55). The formation of free radicals and release of glutamate into the extracellular space are proportional to the intraischemic brain temperature (Fig. 24.12). Hyperthermia stimulates N-methyl-D-aspartate receptors, which increase intracellular calcium levels and enhance the free radicals resulting from the release of arachidonic acid (56,57).
|
|
|
Figure 24.12. Time course of changes in striatal microdialysis-perfusate levels of glutamate in rats subjected to a 20-minute global forebrain ischemia by four-vessel occlusion at intraischemic brain temperatures of 30°C, 33°C, or 36°C. In normothermic rats, a massive extracellular release of glutamate was triggered by the ischemic insult, while in animals with hypothermic brain temperatures (33°C or 30°C), glutamate release was almost entirely suppressed. (From Busto R, Globus MY-T, Dietrich WD, et al. Effect of mild hypothermia on ischemia-induced release of neurotransmitters and free fatty acids in rat brain. Stroke. 1989;20:904–910, with permission.) |
Therapeutic Window
Anoxic brain injury is mainly proportional to the duration of ischemia, with death occurring within about 5 minutes of complete anoxia. Therefore, many physicians believe that hypothermia must be induced as early as possible. In other words, the earlier therapeutic hypothermia is started and the target temperature is reached, the greater the chance of a positive outcome (58,59).
The recommended duration for therapeutic hypothermia is 12 to 24 hours in the postresuscitation period. It is also reported that patients obtain best outcomes when therapeutic hypothermia is maintained between 48 and 72 hours (60,61). There is a consensus—although not evidence based—that patients should be very slowly rewarmed from therapeutic hypothermia.
Fever
Hyperthermia is a generic term simply indicating a core body temperature exceeding normal values. In contrast, fever is a regulated increase in the core temperature targeted by the thermoregulatory system. Hyperthermia can result from a variety of causes and usually indicates a problem of sufficient severity that physician intervention is required. Fever is a special type of regulated hyperthermia that synchronously increases all thermoregulatory response thresholds. Fever thus represents an increase in the body's temperature set point. Fever is common among patients with neurologic injury and is associated with worse outcome from stroke, although fever probably improves infectious outcomes (62,63).
Normal body temperature is neither set nor maintained by circulating factors. In contrast, fever results when endogenous pyrogens increase the thermoregulatory target temperature (“set point”). Identified endogenous pyrogens include interleukin-1, tumor necrosis factor, interferon-α, and macrophage inflammatory protein-1 (64). Although it was initially believed that these factors acted directly on hypothalamic thermoregulatory centers, there is increasing evidence for a more complicated system involving vagal afferents. Most endogenous pyrogens have peripheral actions (e.g., immune system activation) in addition to their central generating capabilities (65,66).
Optimal treatment strategies for fever have yet to be established. Although active surface cooling of febrile patients makes intuitive sense, it is often either ineffective or counterproductive. This is principally because the febrile target set by the hypothalamus is in terms of mean body temperature, not core temperature. Weighted mean body temperature (for thermoregulatory purposes) is a linear combination of core and skin temperatures, with the skin contributing approximately 20% (67). A decreasing skin temperature of 4°C will thus increase the target core temperature by about 1°C. In intact patients, thermoregulatory defenses are more powerful than most types of surface cooling. The result is that active cooling fails to reduce core temperature, but does make patients highly uncomfortable, increases plasma catecholamine concentrations, augments metabolic rate, and provokes hypertension (Fig. 24.13) (68). However, in patients with restricted thermoregulatory defenses, surface cooling is an effective treatment for fever (69,70). The general strategy, then, is to distinguish active fever from passive hyperthermia. In the case of fever, treat the underlying fever source whenever possible because active cooling provokes considerable stress and usually is not particularly effective. In cases where hyperthermia appears passive or results from excessive heat production, simple cooling measures will usually be effective.
|
|
|
Figure 24.13. Change in core and skin temperature and thermal comfort after fever was induced by administration of interleukin-2, starting at elapsed time zero. The designated thermal management started after 3 elapsed hours and continued for 5 hours. This consisted of no treatment (control), self-adjustment per the subjects' comfort (self-adjust), and forced-air cooling (cooling). Thermal comfort is reported as millimeters on a visual analog scale (VAS), with 0 mm indicating the worst imaginable cold, 50 mm identifying thermoneutrality, and 100 mm being the worst imaginable heat. Active cooling made the subjects feel miserably cold, but did not reduce core temperature. Data are presented as means ± standard deviations. (From Lenhardt R, Negishi C, Sessler DI, et al. The effects of physical treatment on induced fever in humans. Am J Med. 1999;106:550–555, with permission.) |
Diet-induced Thermogenesis
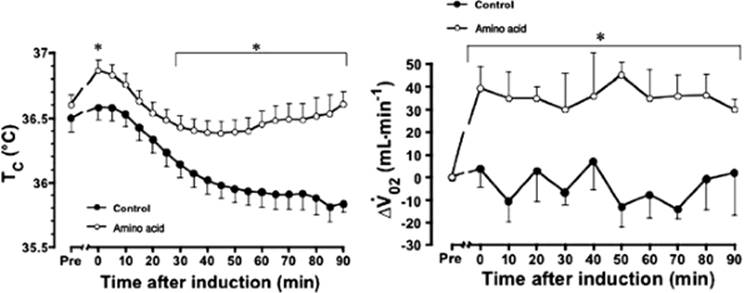
The increase in energy expenditure after the ingestion of a meal or infusion of nutrients has been referred to as dietary-induced or nutrient-induced thermogenesis. The thermogenesis induced by protein or amino acids is known to be largest and most prolonged. An intravenous infusion of 600 kJ (35 g) of amino acids increases energy expenditure in awake, healthy volunteers by 20%, and thus increases temperature of blood in the hepatic artery and vein by 0.3°C (71). In patients under general anesthesia for lower abdominal surgery, the continuous infusion of amino acids (240 kJ/hour) for 2 hours keeps core temperature about 0.3°C/hour greater than in patients not receiving the infusion (Fig. 24.14) (72). Furthermore, a continuous infusion at a rate of 4 kJ/kg/hour, for 2 hours before induction of spinal anesthesia, prevents hypothermia (Fig. 24.15) (73). Recently, a perioperative amino acid infusion at a rate of 4 kJ/kg/hour for 4 hours has been shown to increase the esophageal temperature at the end of surgery by 0.6°C, in addition to shortening the artificial ventilation period, intensive care unit (ICU) stay, and hospital stay after off-pump coronary artery bypass grafting (74).
|
|
|
Figure 24.14. Changes in rectal temperature from baseline measurements throughout anesthesia in eight patients receiving amino acid infusion for 1 hour before and 1 hour of anesthesia, eight patients receiving amino acids for 2 hours before anesthesia, and eight patients receiving a nutrient-free saline solution for 1 hour before and 1 hour of anesthesia. Vertical bars = standard error of mean. (From Sellden E, Branstrom R, Brundin T. Preoperative infusion of amino acids prevent postoperative hypothermia. Br J Anaesth. 1996;76:227–234, with permission.) |
Fructose is also known to elicit the greatest thermogenesis among the various carbohydrates that have been tested. Oral intake of 75 g fructose in awake healthy volunteers increases arterial blood temperature by more than 0.2°C (75). A continuous infusion of fructose, at a rate of 0.5 g/kg/hour for 4 hours in patients during general anesthesia for lower abdominal surgery, increased esophageal temperature at 3 hours after induction by 0.6°C. Fructose also increases the metabolic rate by approximately 20%, which is presumably the mechanism by which it increases body temperature (Fig. 24.16) (76).
Summary
The signal from the cutaneous thermosensors detecting the change in environmental temperature is transmitted to the thermoregulatory center in the hypothalamus, which, in turn, controls efferent defenses. The thermoneutral zone can be defined as the range of environmental temperature over which cutaneous vasomotion alone can maintain body temperature, and ranges from roughly 29°C to 31°C.
Cutaneous vasoconstriction is the first autonomic response to cold. The normal threshold (triggering core temperature) for vasoconstriction is about 36.5°C. Shivering is the final autonomic response that is activated to cold and is generally only observed when behavioral responses and vasoconstriction fail to maintain an adequate core temperature. Vigorous shivering doubles the metabolic rate. The shivering threshold is approximately 35.5°C.
Patients undergoing mild hypothermia exhibit decreased heart rate, increased systemic vascular resistance, normal mean blood pressure, and stroke volume. Minute ventilation decreases during therapeutic hypothermia, but PaCO2 remains normal because of the decreased metabolic rate. The number of leukocytes decreases. Prolonged hypothermia also decreases the number and function of platelets. During induction of therapeutic hypothermia, potassium shifts from the plasma into cells, resulting in hypokalemia. Conversely, during rewarming, the shift of potassium back into the plasma may lead to life-threatening hyperkalemia. Urine output increases at induction of hypothermia. Gut motility decreases during hypothermia, which delays enteral feeding. Blood glucose concentrations increase in response to reduced insulin secretion.
|
|
|
Figure 24.15. Change in tympanic membrane core temperature (Tc) during spinal anesthesia. Tc values at 0 minutes and after 30 minutes were significantly greater in the amino acid infusion group than the saline group. Data are presented as means ± standard error of mean. “Pre” indicates the period of pre–amino acid infusion. Asterisks (*) indicate a statistically significant difference between the groups (P <0.05). (From Kasai T, Nakajima Y, Matsukawa T, et al. Effect of preoperative amino acid infusion on thermoregulatory response during spinal anaesthesia. Br J Anaesth. 2003;90:58–61, with permission.) |
Thermoregulatory control is based on the integrated temperatures from tissues throughout the body, not just core temperature. Skin temperature contributes approximately 20% to thermoregulatory control. Decreasing skin temperature by 4°C will thus increase the target core temperature by about 1°C. In intact patients, thermoregulatory defenses are more powerful than most types of surface cooling. The result is that active cooling fails to reduce core temperature, but does make patients highly uncomfortable, increases plasma catecholamine concentrations, augments metabolic rate, and provokes hypertension. However, in patients with restricted thermoregulatory defenses, surface cooling is an effective treatment for fever.
|
|
|
Figure 24.16. Core temperature measured at the distal esophagus (Tes) during surgery. Patients receiving the fructose infusion had significantly greater core temperatures than those receiving saline (*P = 0.001), starting 20 minutes after induction of anesthesia until the end of the monitoring period (180 minutes after induction). Data are presented as mean ± standard deviation for ten patients in each group. (From Mizobe T, Nakajima Y, Ueno H, et al. Fructose administration increases intraoperative core temperature by augmenting both metabolic rate and the vasoconstriction threshold. Anesthesiology. 2006;104:1124–1130, with permission.) |
Peripheral vasoconstriction plays a major role in the thermoregulatory response to reduced body temperature. The first response to cold is vasoconstriction, which reduces heat loss from the skin surface. Therefore, the change in cutaneous circulation in a nonthermoregulatory reflex, such as the baroreceptor reflex, interacts with the thermoregulatory vasoconstriction.
After the ingestion or infusion of nutrients, there is an increase in energy expenditure, which has been referred to as dietary- or nutrient-induced thermogenesis. This thermic effect of nutrition is divided into an obligatory component, representing the theoretically metabolic costs for the processing and storage of the nutrients ingested, and the facultative component, representing a general stimulation of the whole-body energy expenditure. Perioperative administrations of amino acids or fructose increase energy expenditure and core temperature during surgery.
References
1. Blumenthal I. Fever—concepts old and new. J R Soc Med. 1997;90(7):391–394.
2. Haller JS Jr. Medical thermometry—a short history. West J Med. 1985;142(1):108–116.
3. Ring EF. The historical development of thermometry and thermal imaging in medicine. J Med Eng Technol. 2006;30(4):192–198.
4. Smith LS. Reexamining age, race, site, and thermometer type as variables affecting temperature measurement in adults—a comparison study. BMC Nurs. 2003;2:1.
5. Blumenthal I. Should we ban the mercury thermometer? J Royal Soc Med. 1992;85:553.
6. Daanen HA. Infrared tympanic temperature and ear canal morphology. J Med Eng Technol. 2006;30(4):224–234.
7. Cork RC, Vaughan RW, Humphrey LS. Precision and accuracy of intraoperative temperature monitoring. Anesth Analg. 1983;62(2):211–214.
8. Bissonnette B, Sessler DI, LaFlamme P. Intraoperative temperature monitoring sites in infants and children and the effect of inspired gas warming on esophageal temperature. Anesth Analg. 1989;69(2):192–196.
9. Erickson R. Thermometer placement for oral temperature measurement in febrile adults. Int J Nurs Stud. 1976;13(4):199–208.
10. Heinz J. Validation of sublingual temperatures in patients with nasogastric tubes. Heart Lung. 1985;14(2):128–130.
11. Tandberg D, Sklar D. Effect of tachypnea on the estimation of body temperature by an oral thermometer. N Engl J Med. 1983;308(16):945–946.
12. Ramsey JG, Ralley FE, Whalley DG. Site of temperature monitoring and prediction of afterdrop after open heart surgery. Can Anaesth Soc J. 1985;32:607–615.
13. Benzinger M. Tympanic thermometry in surgery and anesthesia. JAMA. 1969;209(8):1207–1211.
14. Shiraki K, Sagawa S, Tajima F, et al. Independence of brain and tympanic temperatures in an unanesthetized human. J Appl Physiol. 1988;65(1):482–486.
15. Crocker BD, Okumura F, McCuaig DI, et al. Temperature monitoring during general anaesthesia. Br J Anaesth. 1980;52(12):1223–1229.
16. Lilly JK, Boland JP, Zekan S. Urinary bladder temperature monitoring: a new index of body core temperature. Crit Care Med. 1980;8(12):742–744.
17. Satinoff E. Neural organization and evolution of thermal regulation in mammals—several hierarchically arranged integrating systems may have evolved to achieve precise thermoregulation. Science. 1978;201:16–22.
18. Satinoff E, Rutstein J. Behavioral thermoregulation in rats with anterior hypothalamic lesions. J Comp Physiol Psychol. 1970;71:77–82.
19. Kosaka M, Simon E, Walther O-E, et al. Response of respiration to selective heating of the spinal cord below partial transection. Experientia. 1969;25:36–37.
20. Mistlberger T, Rusak B. Mechanisms and models of the circadian time keeping system. In: Kryger MH, Dement WC, eds. Principles and Practice of Sleep Medicine. Philadelphia: WB Saunders; 1989:141–152.
21. Crawford DC, Hicks B, Thompson MJ. Which thermometer? Factors influencing best choice for intermittent clinical temperature assessment. J Med Eng Technol. 2006;30(4):199–211.
22. Lopez M, Sessler DI, Walter K, et al. Rate and gender dependence of the sweating, vasoconstriction, and shivering thresholds in humans. Anesthesiology. 1994;80:780–788.
23. Stephenson LA, Kolka MA. Menstrual cycle phase and time of day alter reference signal controlling arm blood flow and sweating. Am J Physiol. 1985;249:R186–191.
24. Nakajima Y, Mizobe T, Takamata A, et al. Baroreflex modulation of peripheral vasoconstriction during progressive hypothermia in anesthetized humans. Am J Physiol Regul Integr Comp Physiol. 2000;279(4):R1430–1436.
25. Brück K. Thermoregulation: control mechanisms and neural processes. In: Sinclair JC, ed. Temperature Regulation and Energy Metabolism in the Newborn. New York: Grune & Stratton; 1978:157–185.
26. Warren JB. Nitric oxide and human skin blood flow responses to acetylcholine and ultraviolet light. FASEB J. 1994;8:247–251.
27. Hall DM, Buettner GR, Matthes RD, et al. Hyperthermia stimulates nitric oxide formation: electron paramagnetic resonance detection of. NO-heme in blood. J Appl Physiol. 1994;77:548–553.
28. Detry J-MR, Brengelmann GL, Rowell LB, et al. Skin and muscle components of forearm blood flow in directly heated resting man. J Appl Physiol. 1972;32:506–511.
29. Hales JRS. Skin arteriovenous anastomoses, their control and role in thermoregulation. In: Johansen K, Burggren W, eds. Cardiovascular Shunts: Phylogenetic, Ontogenetic and Clinical Aspects. Copenhagen: Munksgaard; 1985:433–451.
30. Sessler DI, McGuire J, Sessler AM. Perioperative thermal insulation. Anesthesiology. 1991;74:875–879.
31. Sessler DI, Moayeri A, Støen R, et al. Thermoregulatory vasoconstriction decreases cutaneous heat loss. Anesthesiology. 1990;73:656–660.
32. Plattner O, Ikeda T, Sessler DI, et al. Postanesthetic vasoconstriction slows postanesthetic peripheral-to-core transfer of cutaneous heat, thereby isolating the core thermal compartment. Anesth Analg. 1997;85:899–906.
33. Greif R, Laciny S, Rajek A, et al. Blood pressure response to thermoregulatory vasoconstriction during isoflurane and desflurane anesthesia. Acta Anaesthesiol Scand. 2003;47:847–852.
34. Israel DJ, Pozos RS. Synchronized slow-amplitude modulations in the electromyograms of shivering muscles. J Appl Physiol. 1989;66:2358–2363.
35. Giesbrecht GG, Sessler DI, Mekjavic IB, et al. Treatment of immersion hypothermia by direct body-to-body contact. J Appl Physiol. 1994;76:2373–2379.
36. Horvath SM, Spurr GB, Hutt BK, et al. Metabolic cost of shivering. J Appl Physiol. 1956;8:595–602.
37. Frank SM, Higgins MS, Fleisher LA, et al. Adrenergic, respiratory, and cardiovascular effects of core cooling in humans. Am J Physiol. 1997;272:R557–R562.
38. Khan F, Spence VA, Belch JJF. Cutaneous vascular responses and thermoregulation in relation to age. Clin Sci. 1992;82:521–528.
39. Vassilieff N, Rosencher N, Sessler DI, et al. The shivering threshold during spinal anesthesia is reduced in the elderly. Anesthesiology. 1995;83:1162–1166.
40. Fahey MR, Sessler DI, Cannon JE, et al. Atracurium, vecuronium, and pancuronium do not alter the minimum alveolar concentration of halothane in humans. Anesthesiology. 1989;71:53–56.
41. Bernard SA, Gray TW, Buist MD, et al. Treatment of comatose survivors of out-of-hospital cardiac arrest with induced hypothermia. N Engl J Med. 2002;346(8):557–563.
42. Osborn JJ. Experimental hypothermia: respiratory and blood pH changes in relation to cardiac function. Am J Physiol. 1953;175:389–398.
43. Mild therapeutic hypothermia to improve the neurologic outcome after cardiac arrest. N Engl J Med. 2002;346(8):549–556.
44. Bernard SA, Buist M. Induced hypothermia in critical care medicine: a review. Crit Care Med. 2003;31(7):2041–2051.
45. Bernard SA, Mac CJB, Buist M. Experience with prolonged induced hypothermia in severe head injury. Crit Care (Lond). 1999;3(6):167–172.
46. Valeri CR, MacGregor H, Cassidy G, et al. Effects of temperature on bleeding time and clotting time in normal male and female volunteers. Crit Care Med. 1995;23(4):698–704.
47. Aibiki M, Kawaguchi S, Maekawa N. Reversible hypophosphatemia during moderate hypothermia therapy for brain-injured patients. Crit Care Med. 2001;29(9):1726–1730.
48. Machida S, Ohta S, Itoh N, et al. (Changes in tissue distribution of potassium during simple hypothermia). Kyobu Geka. 1977;30(5):413–418.
49. Zeiner A, Sunder-Plassmann G, Sterz F, et al. The effect of mild therapeutic hypothermia on renal function after cardiopulmonary resuscitation in men. Resuscitation. 2004;60(3):253–261.
50. Arrowsmith JE, Grocott HP, Reves JG, et al. Central nervous system complications of cardiac surgery. Br J Anaesth. 2000;84(3):378–393.
51. Curry DL, Curry KP. Hypothermia and insulin secretion. Endocrinology. 1970;87(4):750–755.
52. van den Berghe G, Wouters P, Weekers F, et al. Intensive insulin therapy in the critically ill patients. N Engl J Med. 2001;345(19):1359–1367.
53. Negovsky VA. Postresuscitation disease. Crit Care Med. 1988;16(10):942–946.
54. Illievich UM, Zornow MH, Choi KT, et al. Effects of hypothermic metabolic suppression on hippocampal glutamate concentrations after transient global cerebral ischemia. Anesth Analg. 1994;78(5):905–911.
55. Safar PJ. Resuscitation of the ischemic brain. In: Albin MS, ed. Textbook of Neuroanesthesia: With Neurosurgical and Neuroscience Perspectives. New York: McGraw-Hill; 1997:557–593.
56. Baena RC, Busto R, Dietrich WD, et al. Hyperthermia delayed by 24 hours aggravates neuronal damage in rat hippocampus following global ischemia. Neurology. 1997;48(3):768–773.
57. Coimbra CR, Plourde V. Abdominal surgery-induced inhibition of gastric emptying is mediated in part by interleukin-1 beta. Am J Physiol. 1996;270(3 Pt 2):R556–560.
58. Silfvast T, Tiainen M, Poutiainen E, et al. Therapeutic hypothermia after prolonged cardiac arrest due to non-coronary causes. Resuscitation. 2003;57(1):109–112.
59. Tisherman SA. Suspended animation for resuscitation from exsanguinating hemorrhage. Crit Care Med. 2004;32(2 Suppl):S46–50.
60. Alzaga AG, Cerdan M, Varon J. Therapeutic hypothermia. Resuscitation. 2006;70(3):369–380.
61. Nolan JP, Hazinski MF, Steen PA, et al. Controversial topics from the 2005 International Consensus Conference on cardiopulmonary resuscitation and emergency cardiovascular care science with treatment recommendations. Resuscitation. 2005;67(2–3):175–179.
62. Kluger MJ. Is fever beneficial? Yale J Biol Med. 1986;59:89–95.
63. Ginsberg MD, Busto R. Combating hyperthermia in acute stroke: a significant clinical concern. Stroke. 1998;29(2):529–534.
64. Davatelis G, Wolpe SD, Sherry B, et al. Macrophage inflammatory protein-1: a prostaglandin-independent endogenous pyrogen. Science. 1989;243(4894 Pt 1):1066–1068.
65. Blatteis CM. Role of the OVLT in the febrile response to circulating pyrogens. Prog Brain Res. 1992;91:409–412.
66. Blatteis CM, Sehic E. Fever: how may circulating pyrogens signal the brain? News Physiol Sci. 1997;12:1–9.
67. Cheng C, Matsukawa T, Sessler DI, et al. Increasing mean skin temperature linearly reduces the core-temperature thresholds for vasoconstriction and shivering in humans. Anesthesiology. 1995;82:1160–1168.
68. Lenhardt R, Negishi C, Sessler DI, et al. The effects of physical treatment on induced fever in humans. Am J Med. 1999;106:550–555.
69. Manthous CA, Hall JB, Olson D, et al. Effect of cooling on oxygen consumption in febrile critically ill patients. Am J Resp Crit Care Med. 1995;151:10–14.
70. Poblete B, Romand J-A, Pichard C, et al. Metabolic effects of i.v. propacetamol, metamizol or external cooling in critically ill febrile sedated patients. Br J Anaesth. 1997;78:123–127.
71. Brundin T, Wahren J. Effects of i.v. amino acids on human splanchnic and whole body oxygen consumption, blood flow, and blood temperatures. Am J Physiol. 1994;266(3 Pt 1):E396–402.
72. Sellden E, Branstrom R, Brundin T. Preoperative infusion of amino acids prevents postoperative hypothermia. Br J Anaesth. 1996;76(2):227–234.
73. Kasai T, Nakajima Y, Matsukawa T, et al. Effect of preoperative amino acid infusion on thermoregulatory response during spinal anaesthesia. Br J Anaesth. 2003;90(1):58–61.
74. Umenai T, Nakajima Y, Sessler DI, et al. Perioperative amino acid infusion improves recovery and shortens the duration of hospitalization after off-pump coronary artery bypass grafting. Anesth Analg. 2006;103(6):1386–1393.
75. Brundin T, Wahren J. Whole body and splanchnic oxygen consumption and blood flow after oral ingestion of fructose or glucose. Am J Physiol. 1993; 264(4 Pt 1):E504–513.
76. Mizobe T, Nakajima Y, Ueno H, et al. Fructose administration increases intraoperative core temperature by augmenting both metabolic rate and the vasoconstriction threshold. Anesthesiology. 2006;104(6):1124–1130.