Learning objectives
• To describe the physiological processes involved in coitus.
• To describe the morphology and characteristics of the male and female gametes and how they are adapted to their specific function.
• To discuss factors thought to be involved in the conception of either a male or a female baby.
• To describe capacitance, the acrosome reaction and the cortical reaction in the stages of fertilization.
• To describe events in the first week after fertilization: the first mitotic divisions, cell cleavage, compaction, hatching, implantation and maternal recognition of pregnancy.
• To outline the relevance and implications of parental imprinting.
• To outline common causes of infertility.
• To discuss the approaches used in assisted reproduction technologies.
Introduction
Fertilization is a series of processes that culminate with the union of the male gamete (the sperm) and the female gamete (the oocyte) to form a diploid zygote. Following fertilization, one cell progressively divides into six trillion cells (6 × 1012), forming a unique individual in about 38 weeks. Understanding fertilization is important for the following events during pregnancy and understanding some of the causes of infertility and failed pregnancy. Knowledge about fertilization had led to the development of sophisticated methods of in vitro fertilization (IVF) and, conversely, strategies for preventing fertilization (the basis for contraception).
Chapter case study
Zara and James have attended the local hospital for the 12-week scan and have been informed that there is now only one baby present in the uterus. It would appear that one of the sacs, observed at Zara's earlier scans, had failed to develop further and the ultrasound scan could no longer detect any evidence of the sac's remnants. Zara and James are understandably upset over the loss of one of their babies but are reassured that the other baby has an extremely low risk of having Down's syndrome, is growing well and appears normal.
• How would you explain to Zara and James what were the possible reasons for one of the embryos failing to develop and is it more likely to occur in heterozygous or homozygous twins?
• How common is the loss of a twin in early pregnancy and what reasons could explain why this happens?
Mammalian fertilization occurs within the female reproductive tract. Less than one in a million sperm, or spermatozoa, reach the oocyte. The sperm are deposited in the vagina during intercourse (coitus) and then make the long journey through the female reproductive tract (equal to about 10000 times their own length). The hazards and challenges imposed on the successful fertilization are thought to help ensure that the individual sperm that actually fertilizes the oocyte is strong and healthy. There is extensive and intricate crosstalk between the oocyte and the fertilizing sperm, which leads to activation of the egg and sperm head decondensation. Following fertilization, the female and male pronuclei form, syngamy occurs and the zygote undergoes the first cleavage divisions while travelling through the uterine tube to the uterus where it implants in the uterine wall.
Coitus
Coitus in humans lasts on average 4 min, which is quite long compared to our closest animal cousin, the chimpanzee, which averages 8 s (De Waal, 1995). Masters and Johnson (described in Levin, 1998) described a four-phase model for sexual responses in humans. This is known as the EPOR model:
• E: excitement phase, when stimuli increase sexual arousal or tension.
• P: plateau phase, when arousal becomes intense; if the level of stimulation is inadequate, arousal subsides and there is no further progression to the next phase.
• O: orgasmic phase, which is a few seconds of involuntary climax during which sexual tension is relieved, usually accompanied by a wave of profound pleasure.
• R: resolution phase, when sexual arousal is dispersed; in males, it is believed that there is an absolute refractory period in which further sexual arousal and orgasm are impossible.
In the male
The principal event necessary for male sexual activity is acquisition and maintenance of penile erection. This is primarily a vascular phenomenon, initiated by neurological controls and facilitated by appropriate psychological and hormonal components. Initial stimulation of the penis can be both psychogenic (from erotic stimuli and sexual fantasy) and tactile (via touch receptors in the penis and perineum) and necessitates autonomic nervous system activity to coordinate increased blood flow into the vascular tissues. Centrally perceived sensual stimuli are relayed via the spinal thoracolumbar erection centre (T11 through L2), and reflex erections, initiated by tactile stimuli to the genital area, activate a reflex arc involving the sacral erection centre (S2 through S4; Dey Shepherd, 2002). Involuntary non-sexual nocturnal erections occur during rapid eye movement sleep. There are three components of erection: increased arterial flow, relaxation of the sinusoidal spaces and venous constriction. In the excitement phase, increased inflow of blood converts the low-volume, low-pressure vasculature to a large-volume, high-pressure system (Andersson and Wagner, 1995).
The arterioles and arteriovenous shunts dilate so there is an increase in blood flow which engorges the erectile vascular tissues (cavernous and spongiosum bodies, see Chapter 2). The corpus spongiosum does not increase in turgor as much as the two corpora cavernosa so the urethra is not compressed. Blood outflow is occluded by compression and constriction of the veins so the sinusoids (blood-filled spaces) enlarge further. This results in hardening and erection of the penis as the blood volume is increased by about 50%.
The control of an erection is by stimulation of parasympathetic nerves, and probably simultaneous inhibition of sympathetic outflow, which reduces arterial smooth muscle tone causing dilatation and increase in blood flow. Many central and peripheral neurotransmitters are involved in mediating the erectile response. Centrally, dopamine, nitric oxide (NO), oxytocin and adrenocorticotrophin (ACTH)/melanocyte-stimulating hormone (α-MSH) seem to have a facilitatory role; serotonin may be either facilitatory or inhibitory, and enkephalins are inhibitory. Peripherally, the balance between vasoconstrictive factors (such as noradrenaline, endothelin and angiotensin) and vasodilatory factors (such as NO, vasoactive intestinal peptide (VIP) and prostanoids) controls the tone of the smooth muscle of the corpora cavernosa and determines the functional state of the penis. Other neurotransmitters implicated in the erectile response are acetylcholine, endothelins and Rho-kinases. NO is considered the most important factor facilitating vasodilatation, thus maximizing blood flow and penile erection; it promotes the generation of cyclic guanosine monophosphate (cGMP) which increases smooth muscle relaxation and inflow of blood. Detumescence (loss of erection) occurs because NO-induced vasodilation abates as cGMP is broken down predominantly by the intracavernosal cGMPphosphodiesterase (cGMP–PDE5). Sildenafil citrate (Viagra) and related oral therapies for erectile dysfunction (ED), such as vardenafil (Levitra) and tadalafil (Cialis), selectively inhibit the breakdown of cGMP by PDE5. Side effects of these drugs may include headache, flushing, rhinitis, urinary tract infections, visual disturbances, diarrhoea and dyspepsia, and they may be contraindicated for men with heart and various other medical conditions.
Descending pathways can be either excitatory (such as those initiated by the perceived attractiveness of the partner) or inhibitory (such as anxiety or guilt). Testosterone, mediated by oestradiol, is required to maintain intrapenile NO synthase levels, which mediate local vasodilatation by increasing NO production. In hypogonadal men, testosterone therapy can restore libido and erectile function. Other hormones, such as prolactin, adrenal steroids and thyroid hormone, contribute to male sexual functioning.
ED (‘impotence’), defined as the inability to achieve and/or to maintain an erection for long enough to permit satisfactory sexual intercourse, can be caused by abnormalities of circulation or of neural inputs affecting the nervous control of erection. ED can result from psychogenic, organic (neurogenic such as spinal trauma, hormonal, vascular, cavernosal or drug induced) or mixed causes. Most cases of ED have an organic origin, mostly vascular disease. Atherosclerosis results in endothelial damage, cellular migration and smooth muscle proliferation influenced by cytokines, thrombosis, growth factors, reactive-oxygen species (ROS) and metabolic changes. Ageing affects NO production by the endothelium. Smoking is a risk factor for ED because it affects the vascular endothelium and nicotine, both increases sympathetic tone leading to smooth muscle contraction in the cavernosal body and decreases activity of the enzyme nitric oxide synthase. Failure of erection can be caused by damage to the spongy bodies, impaired flow in the vessels supplying the penis, drugs that interfere with neurotransmitter action or psychogenic factors. Local atherosclerosis can affect blood flow as can nicotine, which has vasoconstrictive properties.
ED is usually treated with vacuum devices, intraurethral suppositories, intracavernosal injections of vasoactive drugs (such as prostaglandin E1 (PGE1, alprostadil) and papaverine, which increase cAMP levels) and, more recently, with oral antagonists of cGMP–PDE (see above). (However, note that many men do not like or cannot tolerate PDE inhibitors.) Neurogenic ED can also arise from lesions in the nervous system, such as peripheral nerve damage. At the cellular level, alterations in potassium efflux may lead to a state of hypercontraction and lack of erectile response. ED is a common age-related problem so the incidence of ED will increase as longevity increases; ageing is associated with a decline in testosterone which may compromise the oestrogen–androgen crosstalk involved in the control of erection. Diabetes, particularly if coexisting with obesity, is a risk factor for ED and is related to accelerated atherosclerosis, alterations in erectile tissue, neuropathy and changes in hormone levels (Tamler, 2009). Diabetes-associated changes include smooth muscle degeneration, endothelial cell dysfunction, abnormal collagen deposition and high levels of glycosylated end products which reduce NO levels; these result in impaired relaxation of the corpus cavernosum smooth muscle (Sullivan et al., 2002). Medications for hypertension can contribute to ED; β-blockers can cause ‘dry’ ejaculation which is probably due to retrograde ejaculation because the β-blockers are causing bladder neck relaxation.
ED can also be a manifestation of cardiovascular disease; hyperlipidaemia and hypercholesterolaemia affect the production of NO by the vascular endothelium. ED is a symptom not a disease; it may act as a barometer of cardiovascular health and be the first presenting sign of previously undiagnosed hypertension, atherosclerosis and diabetes. Although the incidence of ED is common and increasing, it is underreported, underdiagnosed and undertreated as many men are reluctant to seek help. The condition can have considerable psychological and social impact on the affected man and his partner and their quality of life, causing depression, anxiety and loss of self-esteem.
As the penis becomes erect, the testes increase their blood volume and are drawn up towards the perineum. The dartos muscle contracts so the scrotal skin thickens and contracts. As stimulation proceeds, the plateau phase of emission occurs, where the muscles of the prostate, vas deferens and seminal vesicle undergo coordinated responses that propel spermatozoa and seminal fluid into the urethra. Ejaculation, the orgasmic phase, is the process of ejecting sperm from the urethra following contraction of the urethra smooth muscle. This is accompanied by contraction of the pelvic floor muscles and the accessory muscles including the vesicular urethral sphincter, which prevents retrograde ejaculation into the bladder. Orgasm, or contraction of the muscles, can occur without ejaculation. Oxytocin has a key role in the regulation of erection and, together with endothelin-1, in coordinated contraction of the epididymis and tubules at orgasm (Filippi et al., 2003); oxytocin responsiveness seems to be mediated by oestrogen. The composition of ejaculate changes because the contractions are sequential and there is relatively little mixing of the components (Table 6.1).
|
Table 6.1 Composition of ejaculate during orgasm |
||
|
Fraction |
Dominant Gland |
Particularly Rich in |
|
Initial |
Prostate |
Acid phosphatase |
|
Mid |
Vas deferens |
Sperm |
|
Late |
Seminal vesicles |
Fructose |
Premature or rapid ejaculation is the most common ejaculatory dysfunction and is usually caused by anxiety or emotional stress (Master and Turek, 2001); it is treated with behavioural therapy, pelvic floor muscle exercises, α-adrenoceptor antagonists (‘sympathetic α-blockers’), topical anaesthetics, tricyclic antidepressants and selective serotonin reuptake inhibitors such as fluoxetine and sertraline. There have also been some clinical trials investigating the use of PDE5 inhibitors in the treatment of rapid ejaculation. Neurological lesions, such as spinal cord damage or damage following injury or surgery to the colon or abdomen, can also cause ejaculatory dysfunction; the sympathetic nerves associated with control of sexual functioning can easily be damaged. Other problems, such as diabetes and multiple sclerosis, can cause ejaculatory problems. Drugs, such as medication for hypertension and the common cold, may cause a lack of emission (deposition of the seminal fluid into the posterior or back part of the urethra), resulting in failure of ejaculation. Bladder neck damage in men who have undergone prostate surgery (particularly, transurethral resection of the prostate) can result in retrograde ejaculation. Bloody ejaculation is usually due to haematospermia, a benign, self-limited condition resulting from inflammation of the seminal vesicles, colon or prostate, which is treatable by antibiotics.
In the female
Women have similar responses in coitus to those of men. Tactile stimulation of the perineal region and the glans clitoris as well as psychogenic stimuli elicit the response. The corpora of the clitoris and the labia undergo vascular engorgement. Increased blood flow to the vagina increases transudation and vaginal lubrication. The vagina increases in width and length and the uterus is elevated upwards, which lifts the cervical os to produce a ‘tenting effect’. At orgasm, vaginal and uterine contractions increase in intensity. During orgasm, women may expel fluid from the urethra; the Skene's glands produce this fluid. Sexual responses in the female tend to be more prolonged. Detumescence of the female organs is similar to detumescence of the penis. Orgasm in women seems to be learnt, whereas in men, it is a reflex action; female orgasm is not essential for pregnancy.
Systemic effects occur in both males and females: heart rate and blood pressure increase, accompanied by peripheral vasodilatation. This is followed by the resolution phase.
The gametes
Male gametes
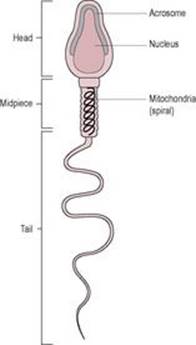
The sperm (Fig. 6.1) develop in the seminiferous tubules of the testes (see Chapter 2). At about 40–50 μm long, of which only approximately 5 μm is the head, the sperm cell is one of the smallest human cells. It retains fertilizing ability for about 2–5 days once deposited in the female reproductive tract. The genetic material of the male gamete is carried in the head of the sperm at the tip of which is located the acrosome, a vesicle containing digestive enzymes. The mid-piece is located between the head and the tail and is packed with mitochondria which generate the ATP required for movement. The tail of the mature sperm has a whip-like action which generates the propulsion for the sperm to swim about 30 cm/h. Abnormalities in human sperm morphology (such as having no tail, two tails or a coiled tail, or no head, two heads or a small head) are very common. Table 6.2 summarizes these and their possible causes. Abnormal motility or morphology of sperm is associated with infertility, causing problems with both fertilization and implantation. As well as the paternal haploid genome (23 paternal chromosomes), the sperm contributes the signal which initiates metabolic activation of the oocyte and also the centriole which causes the microtubules to form the mitotic spindle.
|
|
|
Fig. 6.1 The male gamete: the spermatozoon or sperm. (Reproduced with permission from Brooker, 1998.) |
|
Table 6.2 Abnormalities of sperm |
||
|
Type of Abnormality |
Description |
Possible Causes |
|
Azoospermia (aspermia) |
No sperm present within the ejaculate |
Primary testicular failure; blockage to the vas deferens, that is, infection or trauma |
|
Oligozoospermia (oligospermia) |
Reduced numbers of sperm in the ejaculate (low sperm count) |
Gonadotrophin insufficiency; drugs (social and medical, alcohol, toxins, etc.) |
|
Idiopathic oligospermia |
Low sperm count but physiological parameters normal |
Unexplained |
|
Teratozoospermia (teratospermia) |
Abnormal morphology, for example, giant heads, double tails |
Genetic, toxins, viral infection |
|
Asthenospermia |
Reduced (or lack of) mobility |
Toxins, infection |
|
Sperm agglutination |
Sperm clump together in groups |
Infection, Production of antigens against sperm (autoimmune response) |
Sperm competition
Female choice and male-to-male competition prior to copulation are examples of sexual selection (Lewis et al., 2008). However, it seems that in many species, females typically mate with more than one male at a time when they could conceive. Sperm competition is the competition between sperm from two or more males to fertilize an ovum. There are various mechanisms involved such as sperm number and sperm length, removal of rival sperm, switching off female receptivity to subsequent males and plugging the female reproductive tract. It was proposed that morphologically abnormal human sperm, rather than being due to errors in production, have a role to play in fertilization (Baker and Bellis, 1988). This highly controversial theory suggested that there are two types of sperm: ‘egg-getters’ and ‘blockers’. The morphologically abnormal sperm may be adapted to non-fertilizing roles, particularly preventing passage and successful fertilization by a competitor's sperm. However, the theory has been contested because the metabolic cost of producing non-fertilizing sperm is high and sperm competition is probably better achieved by better swimming and fertilizing ability rather than by producing sperm to compete in other ways (Lewis et al., 2008). Comparative physiology predicts that mammalian species that have a large testis:body weight ratio are more likely to benefit from a promiscuous mating system in which sperm competition operates (because larger testes produce more sperm); by these criteria, men were not designed to be promiscuous (Short, 1997).
Female gametes
The ovum (i.e. the oocyte surrounded by the corona radiata) is expelled from the mature follicle of the ovary (see Chapter 4) and picked up by the fimbria of the uterine tube. The oocyte is the largest human cell, approximately 120–150 μm in diameter, and is therefore just visible to the naked eye. In comparison, follicular cells are typical-sized human cells of about 10 μm diameter. A rim of these follicular cells known as cumulus cells or the corona radiata surrounds the released oocyte. During follicular growth, the oocyte accumulates RNA and protein and numerous large mitochondria, which provide the needs of the dividing zygote. At birth, meiotic division has been suspended (see Chapter 7).
The fertile window
The lifespan of the human oocyte is thought to be about 6–24 h. Sperm have a viability of up to about 5 days in oestrogenized cervical mucus, so sperm in the reproductive tract up to 5 days before ovulation have a chance of fertilizing an oocyte. This means that the ‘fertile window’, when conception is possible, is about 4–5 days before ovulation. Conception on the day after ovulation has never been reported (Stanford et al., 2002). Sperm concentrations fall with increasing frequency of intercourse but not to an extent that daily intercourse reduces benefit. Sperm counts are maximal after about 5 days of abstinence. Thus, daily intercourse during the fertile window optimizes conception because each day of intercourse increases the probability of pregnancy. The best outcome might be achieved by couples abstaining from intercourse for about 5 days prior to the fertile window and then aiming for daily intercourse.
Sex of the zygote
In meiotic division, the cells have their genetic complement reduced from 46 to 23 chromosomes. Each normal sperm will have 22 autosomes and either an X or a Y sex chromosome. If the oocyte is fertilized by a sperm bearing an X chromosome (a gynosperm), the zygote will be female, and if it has a Y chromosome (an androsperm), the offspring will be male (see Chapter 5). Theoretically, there will be equal numbers of gynosperm and androsperm. However, the sex ratio is not constant. The number of male babies born exceeds the number of female babies all over the world and, since the actual number of male conceptions is proportionately higher, male embryos have a higher failure rate (Bromwich, 1991).
It was observed that the incidence of male babies was markedly higher in cultures where menstruating women were considered unclean and had a ritual cleansing period (niddah) before resuming sexual relations (Harlap, 1979). It was therefore suggested that sperm bearing X and Y chromosomes swim at different rates. If there is a longer period between menstruation and the first intercourse, sperm deposition is less likely to occur much before ovulation, so a recently ejaculated sperm will achieve fertilization. It is hypothesized that androsperm, which are slightly smaller than gynosperm and have rounder heads, can swim faster.
There are a number of methods that are proposed to alter the sex chromosome ratio in sperm and allow sex selection. The difference in mass of gynosperm and androsperm in some species is much more marked than it is in humans. Bulls' sperm, for instance, can be effectively separated by differential centrifugation. The bottom fraction in the tube will be enriched with the heavier gynosperm, which can be used to impregnate cows to increase the female dairy proportion of the herd. Separation of human androsperm and gynosperm seems more difficult but is practised albeit with questionable success rates. Some environmental factors may influence the ratio of X- and Y-bearing sperm (James, 2008). Although a number of practices may have no scientific foundation, an increased number of female babies are born as the father ages, infants with blood group O are more likely to be male and environmental pollution seems to increase the number of female babies born. Sex selection is practised but usually not at the time of fertilization. In some parts of the world, selective abortion or infanticide is reported. In the West, sex selection is evident in that couples who have both a male and a female child are less likely to have further children.
Case study 6.1 looks at the question of sex determination.
Case study 6.1
Molly, who has four healthy daughters, is expecting her fifth child. She jokes with the midwife that she knows that this will be a girl as well. The midwife asks her why and Molly informs her that her husband works on an oil rig and is always home for 10 days and then away for 10 days. She laughingly states that every time she has conceived she has always started her period on the day her husband returns home.
• What factors in Molly and her husband's life may influence the sex of their children?
• How unusual is it to have five children all of the same sex?
In the days leading up to ovulation, the epithelial cells lining the uterine tubes become more ciliated and smooth muscle activity of the tubes increases. At ovulation, the fimbriae of the uterine tube move closer to the ovary and rhythmically stroke its surface. These sweeping movements, together with the currents generated by the moving cilia, facilitate the capture of the ovum released at ovulation. Ovum capture is remarkably efficient. Some women have only one functional ovary and one functional uterine tube; even if the ovary and uterine tube are from opposite sides, pregnancy can still occur. The oocyte is transported towards the uterus by movements generated by peristaltic contractions of the uterine tube aided by the beating movement of the cilia. The oocyte has no inherent motility but is washed along by tubal fluid secreted by the epithelial cells and serum transudate. It takes about 3 or 4 days to reach the uterus. Initially, the movement through the ampulla, where fertilization is most likely to occur, is slow, but the zygote travels faster through the isthmus into the uterus. The junction between the uterine tube and the uterus relaxes under the influence of progesterone and allows the oocyte through. If the oocyte has not been fertilized, it degenerates and is phagocytosed.
Stages of fertilization
Gamete motility and sperm deposition
Spermiogenesis (see Chapter 2) occurs in the seminiferous tubules but, although the sperm are morphologically mature, they are not fully motile. The sperm develop swimming ability during a maturation phase of 4–12 days in the epididymis. At intercourse, about 200–300 million sperm are released in about 3 mL of seminal fluid (Box 6.1), which is deposited in the vagina. Repeated ejaculation normally results in a fall in sperm concentration, but the proportion of motile sperm decreases in men who are infertile, suggesting that impaired transport through the male genital tract affects motility (Matilsky et al., 1993). The sperm coagulate in the vagina, which appears to facilitate their retention and to buffer them against the normally unfavourable acidic environment (pH ≈ 4–5) of the vagina. The pH of the vagina is increased by the buffers in the seminal fluid favouring sperm motility and access to the cervix. The coagulum dissolves in about 20–60 min.
Box 6.1
Composition of ejaculate
• 40–250 million sperm
• Prostatic fluid (30%): citric acid, acid phosphatase, magnesium and zinc ions
• Seminal fluid (60%): fructose (energy source for sperm), alkaline
• pH 7.0–8.3
• Volume: 2–6 mL
Between days 9 and 16 of the menstrual cycle, during the fertile period of the few days preceding and including ovulation, the watery composition of cervical mucus facilitates passage of sperm (see Table 4.1, p. 83). The cervical mucus interacts with the sperm and provides protection and nourishment. It also acts as a reservoir and may filter out sperm with abnormal morphology and motility (Suarez and Pacey, 2006). Most sperm (99%) do not enter the uterus. A few hundred sperm reach the uterine tubes within a few hours of coitus; this first wave of rapid transport probably depends on rhythmic muscular contractions of the female reproductive tract. Some sperm appear to be stored in a reservoir (or at least remain in a functional and fertile state associated with the isthmic epithelium) within the uterine tubes for up to 24 h; they are activated by ovulation. Muscular activity of the female genital tract does not seem to be essential for fertilization; some sperm will be stored in cervical crypts and then travel in a relatively slow second wave through the cervical mucus, reaching the uterine tube a few days after ejaculation. Chemoattractants, released from the ovum and the surrounding cumulus cells and temperature changes, guide the sperm to the cumulus cell oocyte complex (Sun et al., 2005).
Capacitation
Ejaculated sperm are unable to fertilize an oocyte immediately; in order to fertilize the oocyte, ejaculated sperm undergo capacitation, interact with the zona pellucida and undergo the acrosome reaction. In vitro, there may be a delay of several hours before unprepared sperm can fertilize an oocyte. However, in vivo (and in sperm reclaimed from the uterus), the action of female enzymes and the oestrogen-stimulated high salt concentration of the uterine secretions speed up the preparation of the sperm. These biochemical and functional changes undergone by the sperm in the uterus and uterine tubes are known as capacitation. The changes include removal of adherent seminal plasma proteins from the sperm, remodelling of the sperm plasma membrane, including changes in cholesterol and phospholipid content, influx of extracellular calcium, increase in cAMP, phosphorylation of proteins and a decrease in intracellular pH (Wassarman, 2009). The modifications alter ion channels in the membrane allowing a transmembrane flux of ions, which direct protein phosphorylation, thereby initiating hyperactivation of the sperm. Sperm metabolism changes from oxidative to glycolytic. The hyperactivated sperm tail movements change to become whiplash-like so the sperm thrusts vigorously forward, moving from the sperm storage reserve in the isthmus of the uterine tube to the ampulla (Suarez, 2008). The accentuated lateral head movements generate a boring action, which aids access through the cumulus cells and ZP to the oocyte. The tail is also involved in sperm movement within the oocyte (Van Blerkom et al., 1995).
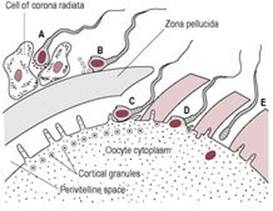
Access to the oocyte
The ovulated oocyte is surrounded by two layers which need to be penetrated by the sperm in order for it to reach the oocyte membrane and to acquire fusibility. The first barrier preventing access of the sperm to the oocyte is the outer layer of cumulus cells, the corona radiata, embedded in an intercellular matrix of carbohydrates, protein and hyaluronic acid. Hyaluronidase, released from the sperm acrosome, breaks down the hyaluronic acid matrix between the follicular cells so sperm can pass through to the zona pellucida. The hyperactive swimming movements of the sperm aid penetration of the corona radiata. The gradual release of sperm from the reservoir of cervical mucus and their activation close to the oocyte means that the time limit of fertility is extended.
Binding to the zona pellucida
The oocyte is surrounded by the zona pellucida, which is about 14–15 μm thick. It is an extracellular matrix composed of sulphated glycoproteins that were produced by the growing oocyte. It is permeable to some viruses, immunoglobulins and enzymes. Before ovulation, cytoplasmic processes from the corona radiata cells penetrate the zona pellucida, allowing communication to, and nourishment of, the oocyte via gap junctions. When these are withdrawn in response to the luteinizing hormone (LH) surge, they may leave gaps in the zona pellucida that offer easier access for sperm penetration, thus facilitating fertilization (Familiari et al., 1992). The zona pellucida acts as a barrier that allows only species-specific sperm–egg interaction.
The human zona pellucida is composed of four glycoproteins ZP1, ZP2, ZP3 and ZP4 (Gupta et al., 2009). ZP2 and ZP3 are implicated as sperm receptors; ZP3 and ZP4 are involved in the binding of sperm to the oocyte and induction of the acrosome reaction. Penetration through the zona pellucida gives the sperm access to the perivitelline space where interaction with the oocyte membrane can take place. The zona pellucida confers a high degree of species specificity; if it is removed, sperm from a different species are able to fertilize an oocyte, though development will rapidly arrest. This is the basis of the hamster zona-free ovum (HZFO) test or sperm penetration assay (SPA) in which hamster oocytes, with their zona chemically removed, are used to test the ability of human sperm to penetrate an oocyte.
The zona pellucida is antigenic: anti-zona pellucida antibodies may be the cause of some cases of infertility. The composition of the zona pellucida changes with the cortical reaction after fertilization so it prevents polyspermy but allows secretions of the uterine tube to reach the oocyte during the early stages of cell division. The zona pellucida also has a role in preventing the blastocyst from prematurely implanting into the wall of the uterine tube before it reaches the uterus. It is possible that an excessively thick zona pellucida could cause problems with blastocyst hatching and subsequent implantation.
Interaction of sperm with the zona pellucida seems to occur in several stages (Ikawa et al., 2010). At first, the capacitated sperm loosely and reversibly adhere to the surface of the zona pellucida. Then, the sperm become strongly and irreversibly bound to the zona pellucida. Many sperm bind to the oocyte zona pellucida but usually only a few sperm penetrate into the perivitelline space and only one will fuse with the oocyte plasma membrane.
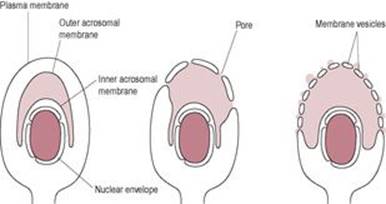
The acrosome reaction
After binding to the zona pellucida, the sperm undergo the acrosome reaction (Fig. 6.2). The sperm binding to the ZP receptor is a carbohydrate-mediated event which initiates a signal transduction cascade and raised intracellular calcium concentration. Carbohydrate-binding proteins on the sperm surface recognize glycoproteins on the ZP. This molecular interaction is associated with inhibition of both the innate and adaptive immune responses (Barroso et al., 2009) which may help to protect the gametes and developing embryo (‘fetoembryonic defence system hypothesis;’Clark et al., 2001). The acrosome reaction can also be triggered by follicular fluid and progesterone (Brucker and Lipford, 1995). The outer acrosome membrane fuses with the covering plasma membrane of the sperm. Small vesicles containing acrosomal enzymes are pinched off and their contents are released. The inner acrosomal membrane is then exposed. A tunnel is digested through the zona pellucida by acrosin, a serine protease that remains bound to the inner acrosomal membrane, and the acrosomal enzymes released from the vesicles. The lurching movements of the sperm propel it forward through the zona pellucida and the perivitelline space so that its head is in contact with the oocyte vitelline (surface) membrane. Penetration through the zona pellucida requires both hyperactivated sperm and lysis of the zona pellucida.
|
|
|
Fig. 6.2 The acrosome reaction. |
Gamete fusion
The acrosome reaction triggers changes in the sperm membrane that allow sperm–oocyte binding and then fusion to occur. The acrosome reaction reveals antigens such as Izumo on the sperm which can interact with receptors on the surface of the oocyte (Ikawa et al., 2010). Similar egg cell surface proteins on the oocyte membrane are recognized by the sperm. The surface of the oocyte is covered with microvilli, except in the region overlying the meiotic spindle. Oocyte microvilli surround the head of the sperm preceding fusion and then the sperm plasma membrane is incorporated into the oocyte membrane. Various docking and recognition molecules on both the sperm head and the oocyte are implicated in sperm–oocyte binding and fusion; these include sperm ADAMs (a family of proteins with a disintegrin and metalloprotease domain) and CD9 and other proteins on oocytes (Kaji and Kudo, 2004). In human fertilization, the sperm tail remains motile and is incorporated into the oocyte (Payne et al., 1997). Paternal mitochondria entering the oocyte are selectively degraded (Kaneda et al., 1995). The zygote and resulting embryo have only maternal mitochondria (Gyllensten et al., 1991); the oocyte seems to lose mitochondrial DNA (mtDNA) with increasing maternal age, a factor that may be important in fertility. One technique to deal with this is transfer of the cytoplasm from a younger woman's oocytes to ‘rescue’ oocytes from older women (Barroso et al., 2009). As well as the male nuclear component, the fertilizing sperm also contribute to the centriole and microtubule-organizing centre, from which the first mitotic spindle will develop (Van Blerkom et al., 1995). Fertilization takes about 10–20 min.
The cortical reaction and block to polyspermy
Polyploidy is usually fatal and is often detected in spontaneously aborted fetuses (Gardner and Evans, 2006). Most human triploidy is due to polyspermic fertilization where two sperm fertilize an ovum. The number of sperm reaching the newly ovulated egg is partly regulated by sperm transit from reservoirs in the female reproductive tract. Oocytes from most mammals develop mechanisms during growth and development to block polyspermy, the entry of more than one sperm. Following fertilization by one sperm, polyspermy is prevented by a sequence of events which modifies the structure of the plasma membrane and/or the zona pellucida. The relative importance to the plasma membrane versus the zona pellucida block to polyspermy varies with species. In most mammalian oocytes, the zona pellucida is thought to provide the most important block to polyspermy. The number of supernumerary sperm found in the perivitelline space between the ZP and the plasma membrane is usually 1–10 in humans, suggesting that the membrane block occurs almost simultaneously with the ZP block. Two events are involved: the cortical reaction and modification of the zona pellucida by enzymes released from the cortical granules (the ‘zona reaction’). Failure of either of these steps results in polyspermy. Polyspermy results in non-diploid zygotes which are usually not viable but, in some cases, can develop into gestational trophoblastic neoplasias and tumours such as the benign hydatidiform mole or the malignant choriocarcinoma (Hauzman and Papp, 2008). The incidence of non-diploid zygotes increases with alcohol, drug use, anaesthesia and fertilization of ‘aged’ oocytes (i.e. aged in terms of hours after ovulation). Intracytoplasmic sperm injection (ICSI) does not trigger the membrane block to polyspermy; this is not important in the clinical scenario, as ICSI-fertilized eggs are not going to be exposed to additional sperm, but it does indicate that ICSI is not fully equivalent to IVF or in vivo fertilization.
The secretion of cortical granules into the perivitelline space is triggered by a rise in intracellular calcium ion (Ca2+) concentration. In experimental species used to study fertilization, such as sea urchin eggs, two mechanisms have evolved to reduce the occurrence of polyspermy. Prior to the ubiquitous zona reaction, sperm binding rapidly triggers an influx of sodium ions, leading to depolarization of the oocyte membrane which transiently blocks binding of additional sperm. This is known as the ‘fast block’ to polyspermy. There is no evidence for membrane depolarization being involved in the fast block to polyspermy in mammals.
The initial calcium increase triggers calcium release from the intracellular stores, which promotes fusion of the cortical granules with the oocyte membrane (Hoodbhoy and Talbot, 1994). The calcium signal starts from the site of fusion and moves as an oscillating wave through the oocyte, sequentially activating about 4000 cortical granules (Fig. 6.3). The trigger for this calcium-induced calcium rise seems to be a soluble cytosolic factor released from the fertilizing sperm. This sperm factor is probably a novel phospholipase C, PLC-ζ (PLC-zeta) which diffuses into the oocyte cytoplasm following fusion of the sperm and oocyte plasma membranes, initiating the calcium release that leads to the oscillations of calcium (Swann et al., 2004).
|
|
|
Fig. 6.3 Fertilization: (A) acrosome reaction; (B) binding to zona pellucida; (C) penetration of zona pellucida; (D) fusion of oocyte and sperm, and cortisol reaction; (E) fertilization. |
These calcium oscillations may last for several hours (Palermo et al., 1997) and act as the signal for the oocyte to activate and begin development. During the ICSI procedure used in IVF (see below), it is common practice to immobilize the sperm by mechanical damage to the sperm tail just before injecting it into the egg (Yanagida et al., 2001). The extensive disruption of the sperm membrane wrought by this damage may help the sperm factor to be released, as there is a more rapid onset of calcium oscillations after ICSI.
Egg activation is triggered by the rise in intracellular calcium concentration; the earliest indicators are the resumption of meiosis and secretion of cortical granules. The rise in calcium subsequently initiates the first division of the embryo (Jones, 2007).
The zona reaction
The contents of the cortical granules (enzymes, such as proteases and peroxidase, and polysaccharides) are released into the perivitelline space and diffuse through the zona pellucida to digest the ZP3 sperm receptors. The zona pellucida loses its ability to bind to sperm and to induce the acrosome reaction. The changed texture of the zona pellucida is described as ‘zona hardening’. This reaction is known as the zona reaction. The composition of the oocyte plasma membrane is also altered.
Events leading to the first mitotic division
Possible benefits of arrested meiosis
Prior to fertilization, the chromosomes of the oocyte had been arrested in metaphase of the second nuclear division (see Fig. 7.11, p 151). It has been suggested that growth of the oocyte requires a diploid number of chromosomes. Meiotic arrest in the diplotene stage of metaphase means that both maternal and paternal alleles can be expressed during oocyte maturation. Observation of IVF has shown that the human meiotic spindle is unstable and very sensitive to external influences.
As a species, humans have a high frequency of aneuploidy in products of fertilization. This results in zygotes and embryos with the wrong number of chromosomes, for example, Down's syndrome with 47 chromosomes (see Table 7.1, p. 152). Down's syndrome is an example of trisomy, a chromosome complement that is compatible with fetal survival; other examples are thought to be just as likely but may result in failed implantation or failed in uterodevelopment. Although chromosome 21 and 22 are both small chromosomes, chromosome 21 has only 225 active genes, whereas chromosome 22 has 545 active genes. Therefore, trisomy 21 results in a relatively small number of genes in triplicate form, which may explain why trisomy 21 is compatible with life. Genes on chromosome 21 are of particular interest because of their role in Down's syndrome, a type of Alzheimer's disease and several types of cancer. These diseases and the small number of genes on chromosome 21 led to it being the second fully sequenced human genes in the Human Genome Project (Hattori et al., 2000).
Aneuploidy is mostly due to non-disjunction of bivalent chromosomes in the first meiotic division (see Fig. 7.11) and is associated with pregnancy loss. Aneuploidy is the most common cause of mental retardation where the fetus survives (Hassold and Hunt, 2001). The frequency of aneuploidy increases with increased maternal age; ageing oocytes seem more prone to errors in meiosis and abnormal segregation of chromosomes. Studies have shown that the risk of Down's syndrome where the extra chromosomal material is maternally derived increases with age. A lower incidence of Down's syndrome results from extra paternal chromosomal material; however, this is less likely to be linked to increasing age of the male partner.
Other factors such as alcohol abuse, chemotherapy and smoking (Zenzes et al., 1995) have been found to disrupt meiosis. Arrest in the second meiotic division may help to prevent aneuploidy and the inclusion of an extra or absent chromosome in the oocyte.
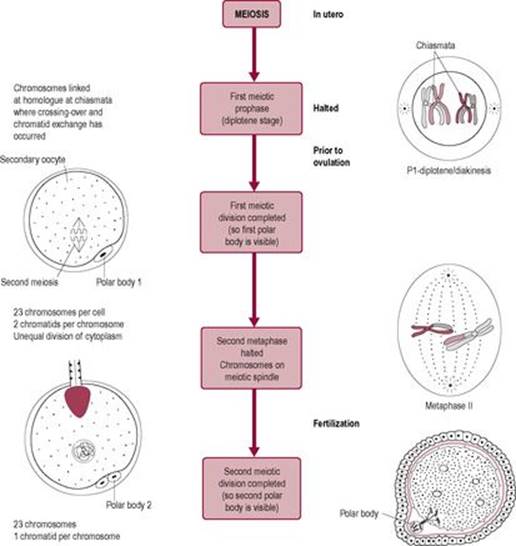
Completion of the second meiotic division
The calcium rise that triggers the cortical reaction is also the stimulus for the oocyte to increase its metabolism. Calcium regulates the cell cycle (Whittaker, 1995); it promotes resumption of meiosis, probably via activation of the M-phase-promoting factor, and completion of second meiotic division (Fig. 6.4). The increase in oxidative metabolism is preceded by a rise in intracellular pH. The 23 paired chromatids then separate, half being expelled as the second polar body into the perivitelline space. A pronuclear membrane appears around the remaining 23 chromosomes forming the female pronucleus, thus completing the meiotic division.
|
|
|
Fig. 6.4 Fertilization and the generation of the second polar body. |
Decondensation of the sperm nucleus
The chromatin or genetic material of the mature sperm is transcriptionally inert and compacted tightly in the head. After the head of the sperm enters the cytoplasm of the oocyte, it is affected by cytoplasmic factors that cause the chromatin threads of the DNA to decondense and become transcriptionally competent. The decondensation takes place while the sperm pronucleus is moving towards the pronucleus of the oocyte. The centrioles radiate microtubules in a formation known as a sperm aster, which aids the movement of the two pronuclei (Schatten, 1994). Then, oocyte histones begin to associate with the male chromosomal material (see Chapter 7). As the developing pronuclei near each other in the centre of the cell, they synthesize DNA in preparation for the first mitotic division and the chromosomes replicate into chromatids.
First mitotic division
In the first mitotic division, the membranes of both pronuclei break down and the male and female chromosomes become organized around a mitotic spindle ready for the first cell division. The combination of the male and female chromosomes is called syngamy. It is at this point that conception has occurred. Fertilization is complete about 18–24 h after fusion, and the fertilized oocyte is known as a zygote (the steps of fertilization are summarized in Box 6.2). A cleavage furrow then appears and the zygote becomes two identical cells.
Box 6.2
Sequence of steps in fertilization
• Deposition of sperm
• Sperm capacitation in the female reproductive tract
• Penetration of the corona radiata
• Binding of capacitated sperm to the zona pellucida
• Acrosome reaction
• Penetration of zona pellucida
• Fusion of sperm with oocyte plasma membrane
• Cortical reaction and prevention of polyspermy
• Increased respiration and metabolism by the oocyte
• Completion of the second meiotic division of the oocyte
• Extrusion of the second polar body
• Decondensation of the sperm nucleus
• Development and fusion of the male and female pronuclei
The random assortment of nuclear material in meiosis and the random exchange of nuclear material create new combinations of genes so the gametes are both haploid and genetically unique. Fertilization of one gamete with another results in a unique combination of genetic material in the zygote, which is important in variation of the species (see Chapter 7). Whether the sperm carries an X or Y chromosome will determine the primary sex of the zygote so it will differentiate into either a female or male embryo (see Chapter 5). Fertilization restores the diploid number of chromosomes and initiates the cleavage of the zygote.
The mitochondria contained in the mid-piece of the sperm enter the egg at fertilization but seldom survive beyond 2 or 3 days after fertilization. The number of mitochondria present in somatic cells ranges from a few hundreds to a few thousands depending on the energy requirements of the specific cell type. Each mitochondrion contains 5–10 identical, circular molecules of mtDNA which codes for 13 proteins involved in the mitochondrial respiratory chain as well as ribosomal RNA (rRNA) and transfer RNA (tRNA) molecules. Mature oocytes are very large cells with high energy requirements and contain approximately 1 million mitochondria. The fertilizing sperm introduces up to 100 mitochondria into the oocyte cytoplasm at fertilization. The sperm mtDNA are highly vulnerable to ROS and mutagenesis during the lengthy process of spermiogenesis, storage, migration through the male and female reproductive tracts and fertilization, particularly as the sperm has little antioxidant capability. So, destruction of potentially defective sperm mitochondria is important to ensure that the developing embryo has a healthy stock of mtDNA. The mechanism by which sperm mitochondria, but not the mitochondria derived from the oocyte, are selectively destroyed relies on the tagging of spermatozoa with ubiquitin during spermatogenesis. Ubiquitin is a small polypeptide universally used by cells to tag protein molecules which are destined for degradation. After initial tagging, sperm mitochondria are subsequently masked by disulphide bond formation during maturation and storage in the epididymis but following fertilization, the glutathione-rich environment of the early embryo exposes the ubiquitin-tagged sperm mid-piece which is then recognized by the oocyte's ubiquitin-proteasome-dependent proteolytic pathway so the sperm mid-piece is lyzed and its component mitochondria and their mtDNA are destroyed. The outcome of this efficient destruction of male-derived mitochondria and their mtDNA is that all mtDNA in human offspring are maternally derived (matrilinear). There is no change in the base pair sequence (of about 16500 base pairs) when the mtDNA is passed on from mother to child so it is a useful tracking tool for tracing ancestry through the female line (Sykes, 2004). The relatively simple analysis of, for example, buccal smears can therefore establish maternal ancestry (see Chapter 7). In forensic investigations, mtDNA rather than nuclear DNA is used to identify human remains such as the remains of the Russian royal family. However, mtDNA has a high mutation rate and aberrant mtDNA are thought to be involved in cancers and disease of ageing such as sporadic Alzheimer's disease (Wallace, 2010).
Development before implantation
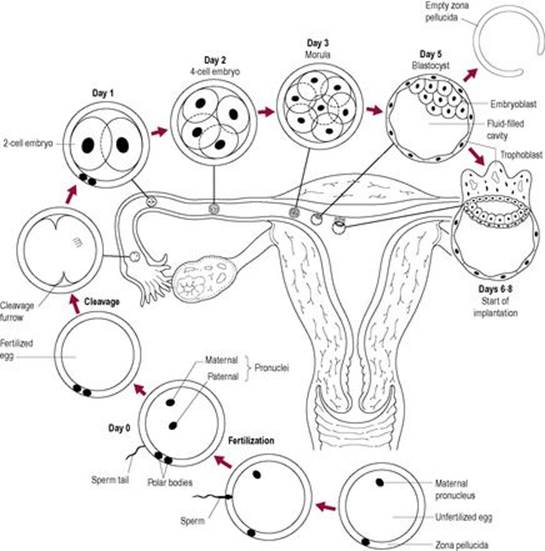
The zygote spends about 4–6 days travelling to the uterus. It is moved through the uterine tube by the peristaltic action of the smooth muscle and the sweeping movements of the cilia and the fluid produced by the ciliated epithelium. The embryo at first divides approximately every 15 h; the division time becomes progressively shorter. During the initial cleavage steps, the embryo is enclosed within the restraining zona pellucida and its total mass remains approximately constant. Cytoplasmic factors regulate cleavage, which occurs without net growth (Fig. 6.5) so cell number increases but the cells become progressively smaller.
|
|
|
Fig. 6.5 Development before implantation. |
Initially, the cleavage divisions are synchronous and each cell (blastomere) is identical, but then the cells divide at an independent rate. Later synchrony is lost, so the pattern of doubling of cell number is also lost. In human and mouse embryos, at the eight-cell stage, each of the cells is totipotent (i.e. is able to generate all cell lineages in the embryo/fetus and extraembryonic tissues) and its fate is not irreversible (Hardy et al., 1990). Experiments on mouse embryos have demonstrated that each of the cells has the capability of independently developing into an embryo. Cells from different origins can also be combined to form a mosaic or a chimera. In human IVF, a single blastomere can be removed from the blastocyst at this stage for genetic testing using the technique of preimplantation genetic diagnosis (PGD) without prejudicing the outcome for the embryo if it is transferred to the uterus (see Chapter 7).
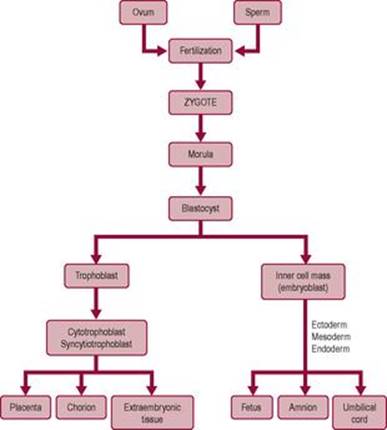
After the eight-cell stage, the cells change morphologically so some of the cells at the outer edge of the embryo become flatter. About day 4, as it reaches the uterus, the embryo is a mass of cells known as the morula (derived from the Latin word for mulberry). The dividing ball of cells enters a phase called compaction. The inner cells are sealed off by outer cells, which adhere tightly in a sphere, developing a polarity and communicating via gap junctions. A fluid-filled cavity, or blastocoele, forms between the inner and outer cell layers; the embryo is now known as a blastocyst. By the 64-cell stage, the cells of the conceptus are irreversibly differentiated on the pathway to becoming embryonic or extraembryonic tissue. Differentiation into a particular cell type seems to be related to positional information of the cells, which induces particular genes to be expressed. Cells of the blastocyst initially differentiate into two distinct cell lines. Most of the outer layer of cells forms the trophoblast, which will develop into the placenta, chorion and extraembryonic tissue. Most of the cells of the inner cell mass will develop into the embryo, umbilical cord and the amnion (Fig. 6.6).
|
|
|
Fig. 6.6 The pathway of cell division. |
Parental imprinting
Chromosomes from the oocyte and sperm express different alleles depending on their origin. Both maternal and paternal chromosomes are required for normal zygote formation and development but genes from both chromosomes are not always expressed. This is described as imprinting and is due to gamete-specific patterns of DNA modification such as methylation. This means that, although the pairs of chromosomes carry the same genetic information, this information is expressed differently depending on whether it originates from the oocyte or the sperm (see Chapter 8). Paternal imprinting favours development of the placenta and inhibits development of the embryo. Maternal imprinting seems to switch off some of the genes involved in placental development. Normally, the paternal copy of the X chromosome is preferentially activated in cells derived from the trophoblast. Conversely, in the inner cell mass, the paternal and maternal chromosomes are randomly inactivated (see Chapter 7). X chromosome inactivation is irreversible, except in oogonia. Parental genetic imprinting is lost (reordered) on formation of the new gametes, which have new parental imprinting on the formation of new chromosome sets. Most imprinted genes are associated with placental tissue and so affect growth and development (Miozzo and Simoni, 2002).
In mouse embryo cells, removal and replacement of one of the pronuclei with another of opposite sex (so both pronuclei are of the same sex) results in abnormal development. If both the pronuclei are female, early embryo development appears normal but placental development is impaired. If both pronuclei are male, placental development appears normal but embryonic development is extremely stunted. Hydatidiform mole development in humans occurs as a result of diandric diploids, that is, when two sperm fertilize an oocyte and the maternal chromosome complement does not participate in development (Jacobs et al., 1980). The result is extreme overdevelopment of the placental tissues and extreme underdevelopment of the embryonic tissues. This situation could also arise from the fertilization of an oocyte with a diploid sperm. Digynic triploids (i.e. two maternal and one paternal chromosome sets) can occur if the polar body is retained.
Human oocytes can be induced to undergo spontaneous cleavage in the absence of fertilization by a sperm and develop into parthenotes. Early stages of cleavage, therefore, seem to be maternally imprinted. However, mouse parthenogenic oocytes arrest after the first cleavage divisions, at the time the embryonic genes would be expressed. Some maternal genes are expressed in the blastocyst but overall, embryonic genes are required for blastocyst formation. Imprinting causes difficulties in cloning such as premature ageing and increased risk of cancer; the clones may not have DNA which is methylated in the correct places. It is also thought that there is an increased risk of low birthweight in otherwise healthy infants born after assisted reproduction technologies (see p. 132), which may be partially related to different conditions very early in life affecting the imprinting of genes involved in fetal growth (Miozzo and Simoni, 2002). Kaguya, a parthenogenic mouse that survived to adulthood (a ‘virgin birth’), has been developed (Kono et al., 2004) by engineering eggs to produce the normally paternally imprinted growth factor, IGF-2. The method was very problematic and only 2 of 457 reconstructed eggs developed; there is a consensus that cloning of humans would be of little merit and be likely to have major risks for maternal health and offspring development (Mollard et al., 2002).
Twins
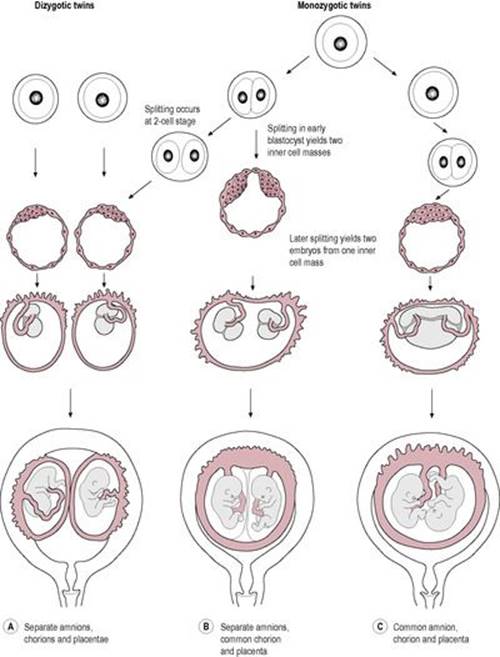
Throughout history, twinning has fascinated civilizations. The incidence of twins in the United Kingdom was about 15 in 1000 livebirths in 2002 (Siddiqui and McEwan, 2007), a increase over previous years because of the increased use of ART, though this figure may drop a little because of the HFEA guidelines to limit the number of embryos transferred during IVF cycles (Milingos and Bhattacharya, 2009). Approximately two-thirds of the twins are dizygotic (fraternal or non-identical) and one-third is monozygotic (identical). The rate of monozygotic twins is fairly constant worldwide but the ratio of monozygotic to dizygotic twins varies markedly from country to country and is affected by season, ethnic origin, parity and maternal age. Dizygotic twins arise from multiple ovulation and two fertilized oocytes implanting and developing; they have separate placentas and membranes (though sometimes these are fused). Monozygotic or identical twins are derived from one fertilized oocyte; the single embryo divides and then splits. Although it is possible that a two-celled embryo could split and result in twins, most twins in practice result from the subdivision of the inner cell mass at the blastocyst stage (Fig. 6.7). About 75% of monozygotic twins share one placenta; the remainder have separate placentas and membranes. Twin pregnancies have a higher rate of obstetric complications, including fetal death, miscarriage and preterm labour, compared to singleton pregnancies.
|
|
|
Fig. 6.7 The pathway of development for dizygotic and monozygotic twins. (Reproduced with permission from Larsen, 1993.) |
Implantation
The blastocyst may remain free-floating in the uterine cavity until implantation at day 7. The blastocyst accumulates fluid and expands. This, together with the digestion and thinning of the zona pellucida by uterine enzymes, results in shedding of the zona pellucida, described as ‘hatching’, at about 6–7 days post-fertilization. The disappearance of the zona pellucida allows the cells of the blastocyst to come into contact with the epithelium of the uterus. The blastocyst consists of at least 100 cells, but some programmed cell death (apoptosis) has already occurred even at this very early stage of the life cycle (Hardy et al., 1989). In most blastocysts, there is a degree of degeneration of some trophoblast cells and some of the inner cell mass. After hatching, growth is no longer physically restricted and the blastocyst grows in mass as well as cell number. Embedding or nidation of the blastocyst normally occurs in the upper part of the body of the uterus (the fundal region). The blastocyst implants at the embryonic pole, where the inner cell mass lies. The inner cell mass forms the embryonic disc (see Chapter 9).
The outer cells of the blastocyst secrete proteolytic enzymes and collagenase, which break down and destroy some of the cells of the endometrial surface, forming a depression in which the blastocyst lies. Implantation in humans is a very invasive mechanism. Uterine muscle activity is low at this time because secretion of progesterone is high. Once implantation has occurred, the lining of the uterus closes over the blastocyst and the pregnancy is established. The trophoblast cells absorb nourishment from the decidua and secrete human chorionic gonadotrophin (hCG) which stimulates growth and secretory activity of the corpus luteum to produce steroid hormones, which support continued growth of the decidua. Critical amounts of hCG are required for blastocyst survival.
Maternal recognition of the pregnancy
Successful implantation requires crosstalk (reciprocated two-way communication between mother and embryo; Hill, 2001). The menstrual cycle, under the control of ovarian steroids, induces the biochemical, physiological and morphological changes which prepare the uterus for blastocyst implantation. Many signals are involved in the crucial role of maintaining the corpus luteum, regulating uterine vascular permeability and maternal immunosuppression. Not all of the signals have been identified. hCG is involved in maintaining the corpus luteum and its essential endocrine role. hCG is luteotrophic, binding to the LH receptors and stimulating a progressive rise in progesterone and oestrogen secretion. Levels of hCG are abnormally low if implantation is inadequate, if trophoblast cell division is insufficient, if implantation is ectopic or if growth of the corpus luteum is deficient (Check et al., 1992). Secretion of hCG from the trophoblast is regulated by trophoblastic gonadotrophin-releasing hormone (GnRH). hCG also binds to the embryonic trophoblast cells and affects the differentiation of cytotrophoblast into syncytiotrophoblast (see Chapter 8). Before it becomes receptive, the uterus contains high levels of the endogenous cannabinoid, anandamine. The concentrations of anandamine fall as receptivity increases, leading to the suggestion that lower birthweight associated with maternal marijuana use occurs because the cannabinoid level is not optimal (Piomelli, 2004).
Implantation and the three stages of apposition, adhesion (attachment) and penetration (invasion) require the endometrium to change and to become responsive. The endometrial stroma is modified (see Chapter 8); vascular permeability increases, and the endometrial cells become hypertrophied and produce prolactin. The processes inducing the changes are known as decidualization; the changed uterine lining is called the decidua. The initial changes begin in the luteal phase of the menstrual cycle. The uterine glands become more tortuous, the spiral arteries develop and the endometrium becomes thicker and oedematous. Uterine secretions increase and intercellular spaces develop. The decidua forms a physical barrier to invasive trophoblast cell migration and generates a cytokine environment that promotes trophoblast attachment rather than invasion (Fazleabas et al., 2004). The decidualized endometrial cells produce IGFBP-1 which modulates interactions at the maternal–fetal interface and is involved in regulating growth.
Effectively, a nidation or implantation window, related to changes in the endometrial epithelium, occurs at about day 20–23 of the cycle (6–8 days after ovulation). The timing of the endometrial changes is mediated by preovulatory increase in 17β-oestriol and subsequent luteal progesterone secretion and has to synchronize with the time the fertilized ovum enters the uterus (Csemiczky et al., 1998). Problems with coincident timing can cause infertility or impaired fertility. During this receptive period, tiny hair-like microvilli protruding from the endometrial cells transiently fuse into single smooth flower-like apical membrane protrusions called pinopodes or uterodomes (Nikas and Aghajanova, 2003). The pinopodes form a week after ovulation and are present for only 2 days before regressing. They absorb fluid and molecules from the uterine lumen which tends to decrease the size of the uterine cavity and to increase the chance of apposition between the embryo and the endometrium. The transient absence of cilia may also prevent the blastocyst being swept away from the site of implantation. Progesterone stimulates pinopode formation, as well as increased uterine secretions, increased blood flow and oedema.
The microenvironment of the reproductive tract is influenced by the secretion of maternal proteins which may then be further modified by proteins secreted by the developing embryo. Embryo-derived platelet-activating factor (ED-PAF) influences vasodilatation and oedema of the decidua. It increases vascular permeability, induces thrombocytopenia, regulates prostaglandin synthesis and activates platelets. Prostaglandins are also involved in regulating decidual factors. Other substances thought to be involved in maternal–conceptus crosstalk include chemokines and cytokines (such as interleukins, interferons and tumour necrosis factor), inhibin and other growth factors (see Box 4.4, p. 74), adhesion molecules (such as integrins), angiogenic and apoptotic factors, leptin and vitamin D (Vigano et al., 2003). The expression of these proteins and peptides can act as a marker of uterine receptivity. The uterine immune system plays an important role in implantation and trophoblastic growth and development; more than 40% of decidua cells are immune cells, predominantly uterine natural killer (NK) cells.
Causes of infertility
Strictly speaking, infertility is a failure to produce gametes capable of fertilization and thus, conception never happens. But in practice, the term infertility is used for failure to continue a pregnancy to term (resulting in a ‘take home’ baby). Some female infertility is due to primary ovarian failure (independent of hypothalamic and pituitary defects) or premature ovarian failure; these probably have genetic components but these have not been clearly identified (Matzuk and Lamb, 2008).
Women who undergo prolonged shock resulting from major haemorrhage (sometimes associated with surgery or a previous pregnancy) can develop anterior pituitary necrosis which can result in Sheehan's syndrome; the pituitary necrosis means that FSH and LH cannot be produced so menopause abruptly begins and fertility is lost.
About a third of female fertility is related to tubal and peritoneal factors (Coughlan and Li, 2009). This includes damage or obstruction to the uterine tubes, often related to pelvic inflammatory disease (PID) or sometimes pelvic or tubal surgery. PID is associated with irreversible damage and loss of the ciliated cells lining the uterine tubes, and thus the loss of transport of the fertilized embryo to the uterus. Persistent tubal damage increases with each episode of infection. Previous ectopic pregnancy, prior abdominal surgery especially if it caused peritubal adhesions, endometriosis, hydrosalpinx (where the uterine tube is blocked and filled with fluid), sepsis following abortion, appendicitis associated with rupture, uterine fibroids and tubal sterilization can also affect tube patency and function. Female fertility can also be curtailed by failure to ovulate or to produce or respond to hormones during the menstrual cycle. Polycystic ovarian syndrome is a relatively common cause of impaired fertility. Ageing affects the success of the menstrual cycle, and it is thought that many women overestimate the length of their reproductive life. Menopause, particularly early or premature menopause, can lead to unexpected early infertility.
In a third to half of infertile couples, the male has poor semen quality which can include low sperm count and/or sperm with abnormal morphology or poor motility (Pacey, 2008). Although male reproductive functions do not cease abruptly in a way equivalent to female menopause, paternal age has marked effects on semen quality, hormone levels, libido and erectile function (Fisch, 2009). Damage to the cells and tissues involved in spermatogenesis such as that caused by trauma or infection (like mumps or malaria) can affect sperm production. Defects of the genital tract (usually obstruction), antibodies or problems with ejaculation usually due to retrograde ejaculation or impotence are the more common causes of male infertility.
There are some factors which can affect fertility in both men and women. These include genetic factors such as Robertsonian translocation (see Chapter 7) where one of the parents has chromosomal rearrangement which means that their own genetic constitution is fine but they have an increased risk that their gametes are unbalanced and could potentially produce a non-viable embryo. Chromosomal abnormalities, especially where the number of sex chromosomes is abnormal, are associated with infertility. Endocrine abnormalities, particularly if they affect the hypothalamic pituitary axis such as pituitary tumours (like prolactinaemia), Kallman's syndrome and hypopituitarism, can affect fertility. Some metabolic/endocrine disorders such as diabetes, liver, kidney, thyroid and adrenal diseases have effects on reproduction. Generally, untreated endocrine disorders are associated with infertility and treatment restores fertility. Sexually transmitted diseases, extremes of body weight in either partner and age can affect fertility. Environmental factors such as virus infections, toxins, pesticides, solvents, pollutants (including cigarette smoke), alcohol and recreational drugs are all associated with reduced fertility (measured as a longer time to conception and/or increased risk of miscarriage). Some medications, particularly those used to treat cancer, such as the chemotherapy regimens, can permanently affect fertility. There is also a significant probability that either both partners have fertility problems or that there is no explanation for failure of conception.
Assisted reproductive technology (artificial fertilization)
Infertility is medically defined at the point when it is appropriate to offer intervention which is when conception has not occurred after 12 months of unprotected intercourse in couples wishing to start a family or after 6 months if the woman is over 35 years old. Subfertility is thought to affect about 15–20% of couples. However, this figure depends on how subfertility is defined as couples may also experience delays in achieving second and subsequent pregnancies. In practice, infertility cannot be cured but subfertility may be treated; medical intervention attempts to optimize conditions that will aid conception. The study of infertility is a relatively new field, open to new research. It involves not just biological aspects of fertility but also social and psychological aspects. Many couples seeking fertility treatment will not succeed. In the United Kingdom, fertility treatment is regulated by the Human Fertilisation and Embryology Act 1990 and monitored by the Human Fertilisation & Embryology Authority (HFEA).
If gametes are available, either from the couple themselves or from a donor, assisted conception techniques can be used. Laboratory techniques can be used to prepare the gametes and bring them closer together to enhance fertility (Box 6.3). Such techniques can be utilized in situations where there is damage to the uterine tubes (the site of normal fertilization), endometriosis, which may alter the uterine environment, male infertility (reduced number, motility or fertilizing ability of sperm) or coital dysfunction. They are also used to treat unexplained fertility where they will test the fertilizing ability of the sperm or where preimplantation diagnosis of genetic disorders is advisable.
Box 6.3
Commonly used assisted reproduction procedures
• In vitro fertilization and embryo transfer (IVF+ET)
• Gamete intrafallopian transfer (GIFT)
• Zygote intrafallopian transfer (ZIFT)
• Intracytoplasmic sperm injection (ICSI)
• Donor insemination (DI)
• Intrauterine insemination (IUI)
Methods of selecting the gametes used for IVF are extremely important especially when legal, ethical or religious reasons mean that destruction of supernumerary embryos has to be avoided. There may be an association between a lower level of ovarian stimulation and better oocyte and embryo quality (Ubaldi and Rienzi, 2008). An oocyte is considered normal when, after the removal of the cumulus cells which are also examined, it has a round clear zona pellucida, a small perivitelline space enclosing a single whole first polar body and cytoplasm which appears pale, moderately granular and without inclusions. However, oocytes with morphological abnormalities can still be fertilized by ICSI, although abnormally shaped oocytes and giant oocytes are not associated with good outcomes. A large perivitelline space may be due to oocyte overmaturity. Likewise, sperm morphology can also be assessed; when strict selection criteria are applied, the success rate is improved and the ICSI technique is known as intracytoplasmic morphologically selected sperm injection (IMSI; Bartoov et al., 2003). The criteria are based on the morphology of the sperm acrosome, neck, mitochondria, tail and nucleus but IMSI is expensive and time-consuming and so is not used routinely.
In vitro fertilization
IVF is now a routine procedure for certain types of infertility. Although IVF has given amazing opportunities to otherwise infertile couples to conceive, there are concerns that the technologies used traverse natural barriers which normally prevent the transmission of genetic defects. There are a number of stages in a cycle of IVF treatment (Box 6.4; Van Steirteghem et al., 1996). The couple may have more than one cause of infertility. IVF can also be used with oocyte donation, for instance, where the woman has ovarian dysfunction (such as premature menopause or Turner's syndrome) or has a high risk of transmitting a serious chromosomal abnormality. Ovarian stimulation is used to increase the number of mature oocytes that will be harvested as some are likely to fail to be fertilized. The method is based on suppressing the natural menstrual cycle by inhibiting the LH surge with a GnRH agonist and then stimulating follicular development with hMG (human menopausal gonadotrophin, which contains FSH and LH), a technique called superovulation. Ultrasound techniques can be used for both follicular assessment and the retrieval of oocytes. When several follicles have grown to a particular size (17–18 mm), ovulation is induced with human gonadotrophin. The oocytes are collected at a preovulatory stage so they are not truly mature. Ovarian hyperstimulation syndrome (OHSS) is a significant complication of fertility treatment involving ovarian stimulation (Nastri et al., 2010); it can cause ovarian enlargement which may result in abdominal distension, increased vascular permeability, intravascular dehydration and the consequences of raised cytokine production such as thrombosis. Progesterone or hCG is necessary to support the luteal phase. The collected oocytes are then cultured. The semen is prepared by separating the motile sperm from the seminal fluid and allowing them to capacitate in an artificial ‘capacitating medium’. The capacitated sperm are introduced to the oocyte within a few hours of oocyte retrieval. The oocyte is enclosed by the corona radiata. The presence of the first polar body shows that the oocyte is at the metaphase II stage (see Fig. 7.11, p. 151) and ready to be fertilized.
Box 6.5.Naegele's rule
This rule was devised by Dr. Franz Karl Naegele who was a German obstetrician in the nineteenth century.
The rule estimates the expected date of delivery (EDD) from the first day of the woman's last menstrual period (LMP) by adding a year, subtracting three months and adding seven days to that date. This approximates to the average normal human pregnancy which lasts 40 weeks (280 days) from the LMP, or 38 weeks (266 days) from the date of fertilization.
Example:
LMP = 8 May 2011
+1 year = 8 May 2012
−3 months = 8 February 2012
+7 days = 15 February 2012
Based on a menstrual cycle of 28 days, add 9 months + 7 days to the first day of the last period to calculate the EDD. Many women have a longer and some have shorter menstrual cycle. If this is the case, ovulation nearly always happens 14 days before period is due – so it is the first part of the menstrual cycle that varies. Therefore, short cycles of 26 days, for instance, need to have another 2 days added on and long cycles, for instance 32 days, need to have four additional days taken off.
It is estimated that only around 5% of babies actually arrive on their due day. Normal term is considered from the completion of the 37th week of pregnancy up until the completion of the 42nd week of pregnancy (often written as T + 14) so under Naegele's rule working from the first day of the last menstrual period, the pregnancy would normally last between 266 and 294 days. After the 41st week, the term ‘post-mature’ is used; current practice is to recommend induction of labour once 42 weeks have been completed. Some babies (around 15%) will be born premature (before the completion of the 37th week).
Box 6.4
Stages in an IVF cycle
• Patient selection
• Ovarian stimulation
• Oocyte retrieval
• Semen preparation
• Possible cryopreservation of gametes (variable success rates)
• Insemination
• Assessment of fertilization
• Embryo cleavage
• Embryo replacement
• Cryopreservation of excess embryos
• Detection of pregnancy
At about 18 h after insemination, the oocytes are denuded by mechanical removal of the cumulus cells. If fertilization has occurred, two distinct pronuclei, and usually two polar bodies, will be observable under a microscope. Twenty-four hours later, embryonic cleavage is evident and the quality of the embryo can be assessed morphologically for its potential to continue developing by the number of anucleate fragments and the evenness of the cells. Embryos with fragmented or uneven cells rarely continue developing. At the four-cell stage, a blastomere can be removed for preimplantation diagnosis (Handyside et al., 1992). Two or three embryos are then selected for transfer to the uterus about 48 h after the two-cell stage; morphological scoring systems are used to assess embryo quality and potential viability. The zona pellucida tends to harden in culture and has to be eroded mechanically or by acid digestion to aid hatching prior to the transfer. Multiple births are common with IVF, although nowadays only three oocytes are implanted into older women, as the success rate is higher in younger women and the protocols try to avoid multiple gestation to increase well-being of the infants. Successful implantation relies on both embryo quality and endometrial receptivity. Excess embryos can be cryopreserved (carefully frozen using protectant fluid and specific rates of freezing and thawing). Increased hCG levels 10–12 days after fertilization confirm a successful pregnancy; levels of hCG usually double every 1.3 days.
In IVF treatment, injections of hCG can be given to prepare the follicles for ovulation following ovarian stimulation as it mimics the action of LH closely. Once the follicles have matured, the oocytes can be harvested. The administration of GnRH agonists administered in the luteal stage of the menstrual cycle has been shown to increase the implantation success of IVF and ICSI treatments (Razieh et al., 2009).
Intracytoplasmic sperm injection
ICSI now accounts for about half of all IVF treatment in the United Kingdom (HFEA, 2009); the other half is conventional IVF. Where there is severe male factor infertility, the sperm are unable to fertilize the oocyte. In ICSI, a single sperm is injected directly into the oocyte cytoplasm (Palermo et al., 1997). The advantage of this method is that it is successful even with very severe sperm dysfunction. Sperm do not need to be motile and can even be harvested from the epididymis by microepididymal sperm aspiration (MESA) or percutaneous epididymal sperm aspiration (PESA). The success rates of ICSI are usually high because the oocyte and endometrium are healthy and apparently not damaged by the procedures (although the cytoplasm of the egg is disrupted during injection and because the hydrolytic enzymes from the sperm acrosome are released into the cytoplasm of the oocyte so there is a small risk of mechanical injury to the meiotic spindle that could potentially lead to aneuploidy). ICSI bypasses almost all of the natural selection mechanisms that challenge the sperm in a normal conception.
It will be important to monitor the health of children born following assisted conception procedures. For instance, the fertility of male offspring resulting from fertilization with a sperm incapable of normal fertilization may be affected. Also, male offspring born following ICSI from fathers with Y chromosome deletions may inherit the same deletion (Chandley, 1998). There are also increased incidences of de novo sex chromosomal aberrations, inheritance of cystic fibrosis mutations and Y microdeletions (Bowdin et al., 2007). Over 1% of births in the United Kingdom and the United States are now due to ART which also accounts for more than 30% of twin births. There is an increased complication rate in infants conceived by IVF, though some of this is probably due to the increased frequency of multiple births. However, singleton IVF infants are more likely to be of lower birthweight and have birth defects. There is an increased risk of the rare growth disorders, such as Beckwith–Weidemann Syndrome and Angelman Syndrome, which are due to defects in gene imprinting (see p. 164). This may be related to the embryo culture conditions used in IVF or the genetic instability underlying the infertility, but the absolute number of children with these disorders is very small.
Cryopreservation techniques have resulted in increased fertility rates; cryopreserved embryos that are surplus to requirement may be offered to other couples who are either infertile or carry a genetic disease but find antenatal diagnosis and termination of an abnormal pregnancy unacceptable. Embryo donation has many ethical, legal and psychosocial implications. There is an increasing trend for patients facing intensive chemotherapy and other procedures to request gamete retrieval and cryopreservation; a more ethically complex area is requests for gamete retrieval from men who are terminally ill, dead, near-dead (for instance, in a persistent vegetative state) or brain-dead.
Case study 6.2 is an example of artificial fertilization.
Key points
• The male gamete (sperm) is one of the smallest human cells and is motile, whereas the female gamete (oocyte) is one of the largest human cells and is immotile.
• Fertilization is the union of the sperm and the oocyte, resulting in a diploid zygote.
• Coitus requires circulatory and neuronal activity resulting in erection of the penis by increasing blood volume. Sexual arousal causes analogous physiological changes in the female.
• Although sex selection is practised in the husbandry of domesticated animals, there is little evidence to suggest that it can be successfully manipulated in humans.
• In order to fertilize the oocyte, sperm undergo a series of changes: (a) development in the testes, (b) maturation in the epididymis and (c) capacitation in the female reproductive tract which alters their tail movements and metabolism and enables them to fertilize the oocyte.
• The acrosome reaction, and release of enzymes, results in digestion of a pathway through the follicular cells and zona pellucida. Interaction between the sperm and the oocyte is mediated by species-specific receptors.
• Fusion of the gametes results in the cortical reaction (which prevents polyspermy), metabolic changes, completion of the oocyte meiotic division, extrusion of the second polar body, DNA replication and initiation of the first mitotic division.
• The embryo or zygote undergoes cell cleavage as it is moved towards the uterus.
• As the number of cells increases, the embryo is described as a morula. The inner and outer cells undergo compaction and differentiate.
• The accumulation of fluid causes the embryo to ‘hatch’ out of the zona pellucida and it is then described as a blastocyst. The blastocyst has an outer layer of trophoblast cells (the future placenta) and an inner cell mass (the future embryo).
• The trophoblast cells secrete hCG, which promotes the survival and further growth of the corpus luteum.
• Maternal recognition of the pregnancy allows embryonic development and uterine receptiveness to be coordinated.
• Implantation occurs about 7 days after fertilization.
• Assisted fertility techniques have been developed for couples who have subfertility.
Application to practice
Primarily, the midwife is not involved with the treatment of infertility. However, with the advance of technology, there are an increasing number of conceptions and successful pregnancies as a result of fertility treatment.
An understanding of the complexities and interventions required to achieve conception is essential in order for the midwife to support women and partners. A knowledge of fertilization is also required in the understanding of contraceptive techniques.
Case study 6.2
Elizabeth is a 42-year-old primipara who has been married for 20 years to Thomas. They had never used contraception. After initial investigations at a subfertility clinic, they were told that Thomas had a very low sperm count and that the sperm present were morphologically abnormal and displayed little motility. After eight attempts of IVF, Elizabeth and Thomas were offered ICSI and as a result, a pregnancy occurred.
• Should the advice that the midwife gives to Elizabeth and Thomas be any different to that given to a fertile couple?
• What kinds of anxieties and concerns would Elizabeth and Thomas have and how might these be addressed?
• Are there any more serious risks and complications for a developing fetus conceived by ICSI?
Annotated further reading
In: (Editor: Balen, A.H.) Infertility in practice ed 3 (2008) Informa Healthcare, London.
A practical guide, based on the authors' clinical practice, which provides an overview of human infertility problems, aetiology and evidence-based possible interventions.
Bancroft, J.H.J., Human sexuality and its problems. ed 3 (2008) Churchill Livingstone, New York .
A comprehensive text of factors determining human sexuality which covers sexual development, anatomy and physiology, biochemistry and endocrinology in addition to sociological and psychological aspects of sexuality. Topics include development of sexual preferences, investigation of sexual dysfunction, marriage, marital therapy, prostitution, AIDS, ageing, sexual abuse and sexual abuse survivors.
Carlson, B.M., Human embryology and developmental biology. ed 4 (2008,) Mosby .
An illustrated textbook detailing the development of the human embryo from conception to birth, including the molecular and mechanistic basis of normal and abnormal development; provides a particularly good description of control of development at the cellular and molecular level.
Coughlan, C.; Ledger, W.L., In-vitro fertilisation, Obstet Gynaecol Reprod Med 18 (2008) 300–306.
A review which succinctly and clearly summarizes the steps involved in in-vitro fertilization.
Devroey, P.; Fauser, B.C.; Diedrich, K., Approaches to improve the diagnosis and management of infertility, Hum Reprod Update 15 (2009) 391–408.
A summary of the current best practice relating to patient pretreatment assessment, ovarian stimulation, embryo assessment and laboratory standards and procedures during ART, together with some brief comments on the future of ART.
Fleming, S.; King, R., Micromanipulation in assisted conception. (2003) Cambridge University Press, Cambridge .
This is a practical handbook which provides comprehensive and well-illustrated descriptions of the micromanipulation techniques used in IVF laboratories, clearly explaining the procedures and potential difficulties.
Furse, A., Your essential infertility companion: a user's guide to tests, technology and therapies. (2009) Thorsons, London .
A useful and sensitively written reference on all aspects of human infertility, including practical information on the medical help available, the procedures involved, potential side effects of fertility drugs, self-help, complementary therapies and emotional aspects of fertility treatment.
Hammarberg, K.; Fisher, J.R.; Wynter, K.H., Psychological and social aspects of pregnancy, childbirth and early parenting after assisted conception: a systematic review, Hum Reprod Update 14 (2008) 395–414.
A review on the interesting and emerging topic of how ART affects parenting and the development of a confident parental identity.
Johnson, M.H., Essential reproduction. ed 6 (2007) Blackwell Science, Oxford .
An excellent, well-organized research-based textbook which explores comparative reproductive physiology of mammals, including anatomy, physiology, endocrinology, genetics and behavioural studies.
Matzuk, M.M.; Lamb, D.J., The biology of infertility: research advances and clinical challenges, Nat Med 14 (2008) 1197–1213.
A detailed review presenting a summary of the current state of knowledge about infertility, including the mechanisms and gene mutations involved.
Sauer, M.V.; Kavic, S.M., Oocyte and embryo donation 2006: reviewing two decades of innovation and controversy, Reprod Biomed Online 12 (2006) 153–162.
An overview of the use of oocyte and embryo donation which discusses reasons for their use, screening of donors and recipients, legal and psychosocial issues and some of the wider implications.
Siddiqui, F.; McEwan, A., Twins, Obstet Gynaecol Reprod Med 17 (2007) 289–295.
A brief description of the causes and management of obstetric complications in twin pregnancies, including in utero death of one twin, discordant growth and anomalies, twin reversed arterial perfusion and twin-to-twin transfusion syndrome.
References
Andersson, K.E.; Wagner, G., Physiology of penile erection, Physiol Rev 75 (1995) 191–236.
Baker, R.R.; Bellis, M.A., Kamikaze sperm in mammals, Anim Behav 36 (1988) 936–939.
Barroso, G.; Valdespin, C.; Vega, E.; et al., Developmental sperm contributions: fertilization and beyond, Fertil Steril 92 (2009) 835–848.
Bartoov, B.; Berkovitz, A.; Eltes, F.; et al., Pregnancy rates are higher with intracytoplasmic morphologically selected sperm injection than with conventional intracytoplasmic injection, Fertil Steril 80 (2003) 1413–1419.
Bowdin, S.; Allen, C.; Kirby, G.; et al., A survey of assisted reproductive technology births and imprinting disorders, Hum Reprod 22 (2007) 3237–3240.
Bromwich, P., The sex ratio, and ways of manipulating it, Prog Obstet Gynaecol 7 (1991) 217–231.
Brooker, C.G., In: Human structure and function ed 2 (1998) Mosby, St Louis, p. 477.
Brucker, C.; Lipford, G.B., The human sperm acrosome reaction: physiology and regulatory mechanisms, Hum Reprod Update 1 (1995) 51–62.
Chandley, A.C., Chromosome anomalies and Y chromosome microdeletions as causal factors in male infertility, Human Reprod 13 (Suppl 1) (1998) 45–50.
Check, J.H.; Weiss, R.M.; Lurie, D., Analysis of serum human chorionic-gonadotropin levels in normal singleton, multiple and abnormal pregnancies, Human Reprod 7 (1992) 1176–1180.
Clark, G.F.; Dell, A.; Morris, H.R.; et al., The species recognition system: a new corollary for the human fetoembryonic defense system hypothesis, Cells Tissues Organs 168 (2001) 113–121.
Coughlan, C.; Li, T.C., Surgical management of tubal disease and infertility, Obstet Gynaecol Reprod Med 19 (2009) 98–105.
Csemiczky, G.; Wramsby, H.; Johannisson, E.; et al., Importance of endometrial quality in women with tubal infertility during a natural menstrual cycle for the outcome of IVF treatment, J Assist Reprod Genet 15 (1998) 55–61.
De Waal, F.B.M., Bonobo sex and society, Sci Am 272 (3) (1995) 82–88.
Dey, J.; Shepherd, M.D., Evaluation and treatment of erectile dysfunction in men with diabetes mellitus, Mayo Clin Proc 77 (2002) 276–282.
Familiari, G.; Notola, S.A.; Macchiarell, G.; et al., Human zona pellucida during in vitro fertilization: an ultrastructural study using saponin, ruthenium red and osmium thiocarbohydrazide, Mol Reprod Dev32 (1992) 51–61.
Fazleabas, A.T.; Kim, J.J.; Strakova, Z., Implantation: embryonic signals and the modulation of the uterine environment – a review, Placenta 25 (2004) S26–S31.
Filippi, S.; Vignozzi, L.; Vannelli, G.B.; et al., Role of oxytocin in the ejaculatory process, J Endocrinol Invest 26 (Suppl 3) (2003) 82–86.
Fisch, H., Older men are having children, but the reality of a male biological clock makes this trend worrisome, Geriatrics 64 (2009) 14–17.
Gardner, A.J.; Evans, J.P., Mammalian membrane block to polyspermy: new insights into how mammalian eggs prevent fertilisation by multiple sperm, Reprod Fertil Dev 18 (2006) 53–61.
Gupta, S.K.; Bansal, P.; Ganguly, A.; et al., Human zona pellucida glycoproteins: functional relevance during fertilization, J Reprod Immunol 83 (2009) 50–55.
Gyllensten, U.; Wharton, D.; Josefsson, A.; et al., Paternal inheritance of mitochondrial DNA in mice, Nature 352 (1991) 255–257.
Handyside, A.H.; Lesko, J.G.; Tarin, J.J.; et al., Birth of a normal girl after in vitro fertilisation and preimplantation diagnostic testing for cystic fibrosis, N Engl J Med 327 (1992) 905–909.
Hardy, K.; Handyside, A.H.; Winston, R.M.L., The human blastocyst-cell number, death and allocation during late preimplantation development in vitro, Development 107 (1989) 597.
Hardy, K.; Martin, K.L.; Leese, H.J.; et al., Human preimplantation development in vitro is not adversely affected by biopsy at the 8-cell stage, Hum Reprod 5 (1990) 708–714.
Harlap, S., Gender of infants conceived on different days of the menstrual cycle, N Engl J Med 291 (1979) 1445–1448.
Hattori, M.; et al., The DNA sequence of human chromosome 21, Nature 405 (2000) 311–319.
Hassold, T.; Hunt, P., To err meiotically is human: the genesis of human aneuploidy, Nat Rev Genet 2 (2001) 280–291.
Hauzman, E.E.; Papp, Z., Conception without the development of a human being, J Perinat Med 36 (2008) 175–177.
Hill, J.A., Maternal–embryonic cross-talk, Ann N Y Acad Sci 943 (2001) 17–25.
Hoodbhoy, T.; Talbot, R., Mammalian cortical granules: contents, fate and function, Mol Reprod Dev 39 (1994) 439–448.
Human Fertilisation Embryology Authority, Fertility facts and figures 2007. (2009) HFEA, London .
Ikawa, M.; Inoue, N.; Benham, A.M.; et al., Fertilization: a sperm's journey to and interaction with the oocyte, J Clin Invest 120 (2010) 984–994.
Jacobs, P.A.; Wilson, C.M.; Sprenkle, J.A.; et al., Mechanism of origin of complete hydatidiform mole, Nature 286 (1980) 714–717.
James, W.H., Evidence that mammalian sex ratios at birth are partially controlled by parental hormone levels around the time of conception, J Endocrinol 198 (2008) 3–15.
Jones, K.T., Intracellular calcium in the fertilization and development of mammalian eggs, Clin Exp Pharmacol Physiol 34 (2007) 1084–1089.
Kaji, K.; Kudo, A., The mechanism of sperm–oocyte fusion in mammals, Reproduction 127 (2004) 423–429.
Kaneda, H.; Hayashi, J.L.; Takahama, S.; et al., Elimination of paternal mitochondrial DNA in intraspecific crosses during early mouse embryogenesis, Proc Natl Acad Sci USA 92 (1995) 4542–4546.
Kono, T.; Obata, Y.; Wu, Q.; et al., Birth of parthenogenetic mice that can develop to adulthood, Nature 428 (2004) 860–864.
Larsen, W.J., In: Human embryology ed 2 (1993) Churchill Livingstone, New York, p. 481.
Levin, R.J., Sex and the human female reproductive tract: what really happens during and after coitus, Int J Impot Res 10 (Suppl 1) (1998) 514–521.
Lewis, Z.; Price, T.A.; Wedell, N., Sperm competition, immunity, selfish genes and cancer, Cell Mol Life Sci 65 (2008) 3241–3254.
Matilsky, M.; Battino, S.; Ben-Ami, M.; et al., The effect of ejaculatory frequency on semen characteristics of normozoospermic and oligozoospermic men from an infertile population, Hum Reprod 8 (1993) 71–73.
Master, V.A.; Turek, P.J., Ejaculatory physiology and dysfunction, Urol Clin North Am 28 (2001) 363–375.
Matzuk, M.M.; Lamb, D.J., The biology of infertility: research advances and clinical challenges, Nat Med 14 (2008) 1197–1213.
Miozzo, M.; Simoni, G., The role of imprinted genes in fetal growth, Biol Neonate 81 (2002) 217–228.
Mollard, R.; Denham, M.; Trounson, A., Technical advances and pitfalls on the way to human cloning, Differentiation 70 (2002) 1–9.
Nastri, C.O.; Ferriani, R.A.; Rocha, I.A.; et al., Ovarian hyperstimulation syndrome: pathophysiology and prevention, J Assist Reprod Genet 27 (2010) 121–128.
Nikas, G.; Aghajanova, L., Endometrial pinopods: some more understanding on human implantation? Reprod Biomed Online 4 (2003) 18–23.
Pacey, A.A., Male fertility and infertility, Obstet Gynaecol Reprod Med 19 (2008) 42–47.
Palermo, G.P.; Avrech, O.M.; Colombero, L.T.; et al., Human sperm cytosolic factor triggers Ca2+ oscillations and overcomes activation failure of mammalian oocytes, Mol Hum Reprod 3 (1997) 367–374.
Payne, D.; Flaherty, S.P.; Barry, M.F.; et al., Preliminary observations on polar body extrusion and pronuclear formation in human oocytes using time-lapse video cinematography, Hum Reprod 12 (1997) 532–541.
Piomelli, D., THC: moderation during implantation, Nat Med 10 (2004) 19–20.
Razieh, D.F.; Maryam, A.R.; Nasim, T., Beneficial effect of luteal-phase gonadotropin-releasing hormone agonist administration on implantation rate after intracytoplasmic sperm injection, Taiwan J Obstet Gynecol 48 (3) (2009) 245–248.
Schatten, G., The centrosome and its mode of inheritance; the reduction of the centrosome during gametogenesis and its restoration during fertility, Dev Biol 165 (1994) 299–335.
Short, R.V., The testis: the witness of the mating system, the site of mutation and the engine of desire, Acta Paediatr Suppl 422 (1997) 3–7.
Stanford, J.B.; White, G.L.; Hatasaka, H., Timing intercourse to achieve pregnancy: current evidence, Obstet Gynecol 100 (2002) 1333–1341.
Sullivan, M.E.; Keoghane, S.R.; Miller, M.A., Vascular risk factors and erectile dysfunction, Br J Urol Int 87 (2002) 838–845.
Suarez, S.S., Control of hyperactivation in sperm, Hum Reprod Update 14 (2008) 647–657.
Suarez, S.S.; Pacey, A.A., Sperm transport in the female reproductive tract, Hum Reprod Update 12 (2006) 23–37.
Sun, F.; Bahat, A.; Gakamsky, A.; et al., Human sperm chemotaxis: both the oocyte and its surrounding cumulus cells secrete sperm chemoattractants, Human Reproduction 20 (2005) 761–767.
Swann, K.; Larman, M.G.; Saunders, C.M.; et al., The cytosolic sperm factor that triggers Ca2+ oscillations and egg activation in mammals is a novel phospholipase C: PLCzeta, Reproduction 127 (2004) 431–439.
Sykes, B., The seven daughters of Eve. (2008) Corgi Books, London .
Tamler, R., Diabetes, obesity, and erectile dysfunction, Gend Med 6 (Suppl 1) (2009) 4–16.
Ubaldi, F.; Rienzi, L., Morphological selection of gametes, Placenta 29 (Suppl B) (2008) 115–212.
Van Blerkom, J.; Davis, P.; Merriam, J.; et al., Nuclear and cytoplasmic dynamics of sperm penetration, pronuclear formation and microtubule organization during fertilization and early preimplantation in the human, Hum Reprod Update 1 (1995) 429–461.
Van Steirteghem, A.; Liebaers, I.; Devroey, P., Assisted reproduction, In: (Editors: Hillier, S.G.; Kitchener, H.C.; Neilson, J.P.) Scientific essentials of reproductive medicine (1996) WB Saunders, Philadelphia, pp. 230–241.
Vigano, P.; Mangioni, S.; Pompei, F.; et al., Maternal–conceptus cross talk: a review, Placenta 24 (2003) S56–S61.
Wallace, D.C., Mitochondrial DNA mutations in disease and aging, Environ Mol Mutagen 51 (2010) 440–450.
Wassarman, P.M., Mammalian fertilization: the strange case of sperm protein 56, Bioessays 31 (2009) 153–158.
Whittaker, M., Regulation of the cell division cycle by inositol phosphate and the calcium signalling pathway, Adv Second Messenger Phosphoprotein Res 30 (1995) 299–310.
Yanagida, K.; Katayose, H.; Hirata, S.; et al., Influence of sperm immobilization on onset of Ca(2() oscillations after ICSI, Hum Reprod 16 (1) (2001) 148–152.
Zenzes, M.T.; Wang, P.; Casper, R.F., Cigarette smoking may affect meiotic maturation of human oocytes, Hum Reprod 10 (1995) 3213–3217.