Frixos Paraskevas
![]() Lymphocytes
Lymphocytes
In the early part of the 20th century, the main debate about the nature and function of the lymphocyte centered around its relationship to cells of the connective tissue and other inflammatory exudate cells. Lymphocytes were considered capable of transforming into granulocytes, monocytes, macrophages, fibroblasts, and other cells. Marschalko (1), Downey (2), Maximow (3), and Bloom (4) all believed that lymphocytes also gave rise to plasma cells. Scientific evidence for these assumptions was lacking, however, and in 1936, Rich said, “The complete ignorance of the function of this cell is one of the most humiliating and disgraceful gaps in all medical knowledge” (5). The first conclusive evidence that antibodies were formed within lymph nodes (LNs) was presented in 1935 by McMaster and Hudack (6), but it took another decade to prove conclusively that lymphoid cells within LNs were the cells that contained antibody (7,8). This discovery tipped the balance in favor of the lymphocytic theory of antibody production (9). In parallel with these studies, the classic work of Fagraeus (10) brought strong indirect evidence of the relationship of plasma cells to antibody production, and the conclusive demonstration by Coons et al. of the presence of antibodies within plasma cells by means of the new and powerful immunofluorescence technique is a landmark in the long debate on the origin of antibodies and the function of plasma cells (11,12). The only remaining question concerning whether plasma cells arise from lymphocytes was answered 10 years later by the elegant studies of Harris et al., in which they demonstrated indisputably the gradual change of a lymphocyte to an antibody-forming plasma cell by means of ultrastructural studies (13,14,15). At the same time, Nowell dispelled the myth that small lymphocytes are end-stage cells incapable of proliferating and taking on new functions (16).
The lymphocytes consist of heterogeneous populations of cells that differ greatly from each other in terms of origin, lifespan, preferred areas of settlement within the lymphoid organs, surface structure, and function. Although some morphologic characteristics, such as size, granularity, and nucleocytoplasmic ratio, distinguish lymphocyte populations from each other, they provide no clues to their lineage and function. The most important, precise, and quantitative method in use in clinical laboratories is based on the identification of certain glycoproteins displayed on the membrane of lymphocytes and collectively called markers. Two remarkable discoveries have helped in the dissemination of routine application of marker analysis in clinical medicine: The development of monoclonal antibodies and the invention of flow cytometry (see Chapter 2).
![]() Morphology of Lymphocytes
Morphology of Lymphocytes
Light Microscopy
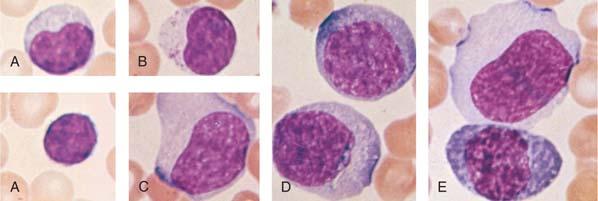
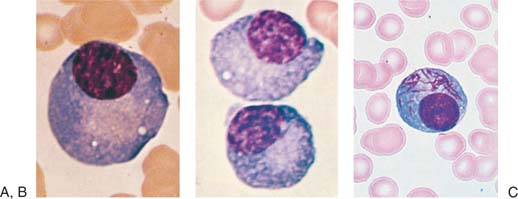
Most lymphocytes in the blood are small (10 μm or smaller), although larger forms are common (Fig. 14.1A). Some of these large lymphocytes are known as large granular lymphocytes because they contain azurophilic granules in their cytoplasm (Fig. 14.1B) (17,18). Often, the distribution of lymphocyte diameters does not yield the commonly described three peaks of small, medium, and large lymphocytes because the dimensions of the cells vary according to the method of preparation. In blood smears stained with Romanowsky dyes, the nucleus is deep purplish blue, usually round or slightly indented, and composed of dense aggregates of chromatin. Nucleoli are not visible with ordinary techniques, although a nucleolus may be seen in wet smears and histologic sections. The cytoplasm forms a narrow rim in small lymphocytes, but it may be abundant in larger cells. The cytoplasm is moderately basophilic and usually is devoid of granules, but larger cells may contain several bright reddish-violet (azurophilic) granules that differ from the granules of myeloid cells in that they do not give the oxidase or peroxidase reaction.
By phase contrast microscopic analysis, a well-defined centrosome can be observed. This area of the cytoplasm is adjacent to the nucleus, and because it is somewhat rigid, it may cause an indentation in the nucleus.
Often, transitional forms between lymphocytes and plasma cells are seen in the blood of patients with viral infections (Fig. 14.1C-E). These cells are variously known as lymphocytoid plasma cells or plasmacytoid lymphocytes. They obviously represent morphologic stages of differentiation of antigenically stimulated lymphocytes.
Transmission Electron Microscopy
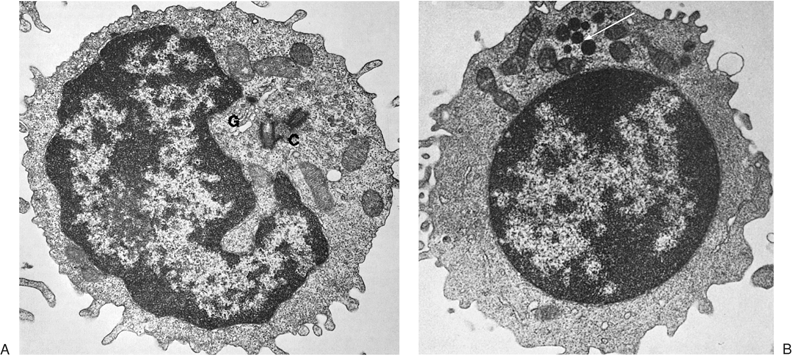
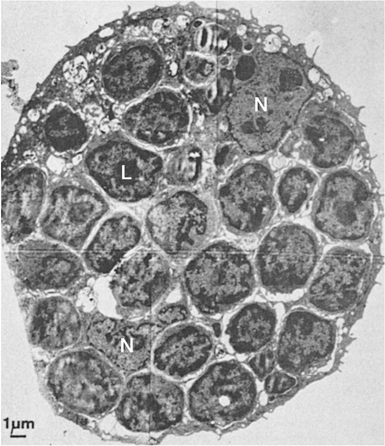
The small lymphocyte (6 to 9 μm in size) reveals a smooth, bi-laminar cytoplasmic membrane that contains only a few microvilli, except in the area of the uropod of motile cells (Fig. 14.2) (19). Occasional blebs, consisting of membrane-bound cytoplasm, are seen on the cell surface and may well be artifacts. The scanty cytoplasm of small lymphocytes shows a remarkable absence of organelles. The Golgi apparatus is small and usually is found near the nuclear notch. One or two centrioles are often seen in the appropriate plane of section. No organized endoplasmic reticulum is observed, although careful scrutiny may reveal one or two profiles. Many free ribosomes and occasional ribosome clusters are evident. Typical mitochondria are common, but lysosomes containing enzymes characteristic of these organelles are sparse. Dense bodies of unknown significance may also be seen occasionally. The cytoskeleton consists of occasional microtubules in the cytoplasm and microfilaments located adjacent to the cell membrane.
|
|
|
Figure 14.1. Morphologic heterogeneity of human peripheral blood lymphocytes. A: Giemsa-stained blood smears: Small and large lymphocytes. B: Large granular lymphocyte with azurophilic granules. C: Atypical lymphocyte. D: Lymphocytes resembling plasma cells (plasmacytoid) from the blood of a patient with viral pneumonia. E: One atypical lymphocyte and one plasmacytoid lymphocyte from peripheral blood. |
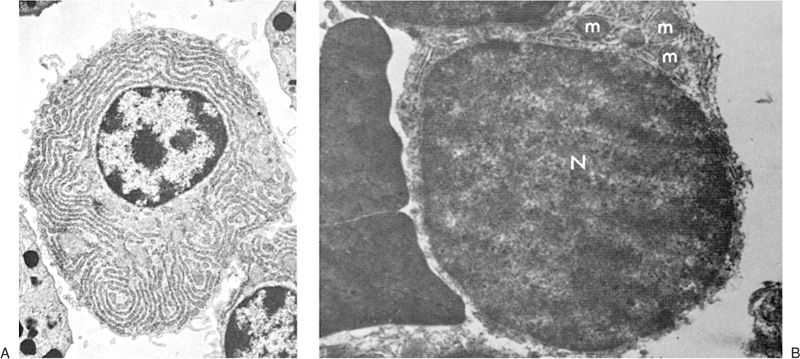
The nucleus is enveloped by a double membrane that fuses at the site of nuclear pores. Abundant dense heterochromatin forms aggregates close to the membrane and less often within the body of the nucleus. These aggregates are separated by interchromatinic spaces containing chromatin, small bits of aggregated chromatin, ribosomelike particles, and fibrils. A nucleolus usually is seen. The medium lymphocyte is larger (6 to 8 μm) because of an increase in the amount of cytoplasm. The Golgi apparatus is more developed than in the small lymphocyte. The cytoplasm also contains numerous polyribosomes and a few strands of endoplasmic reticulum, mostly parallel with the nuclear membrane.
Lymphoblasts usually are larger cells (8 to 12 μm) with loose nuclear chromatin and a giant nucleolus that has a reticulated appearance. It occupies as much as one third of the nuclear area. The cytoplasm contains many polyribosomes, but cisternae of endoplasmic reticulum are scarce. Lymphocytes carry the normal diploid number of 44 autosomes and two sex chromosomes.
|
|
|
Figure 14.2. Ultrastructure of normal peripheral blood lymphocytes. A: A typical resting lymphocyte. The nucleus contains primarily heterochromatin in aggregates along the nuclear membrane. The nucleolus in lymphocytes is usually small. In the cytoplasm, the ribosomes are dispersed, but occasionally, a few short strands of rough endoplasmic reticulum are visible. The mitochondria are well developed and the centrioles (C), longitudinally sectioned in this illustration, show evidence that the cell is ready to enter mitosis on triggering. The Golgi apparatus (G) is small (×24,000). B: This lymphocyte contains granules (arrow) and is likely to correspond to the large granular lymphocyte variety. The cytoplasm is abundant and filled with ribosomes with a few strands of endoplasmic reticulum (×12,000). (From Zucker-Franklin D, Greaves MF, Grossi CF. Atlas of blood cells, 2nd ed. Philadelphia: Lea & Febiger, 1988, with permission.) |
Scanning Electron Microscopy
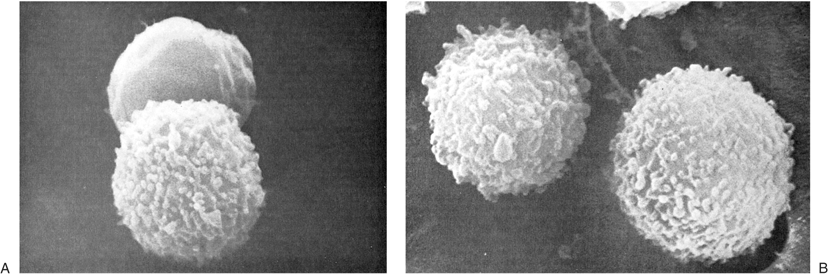
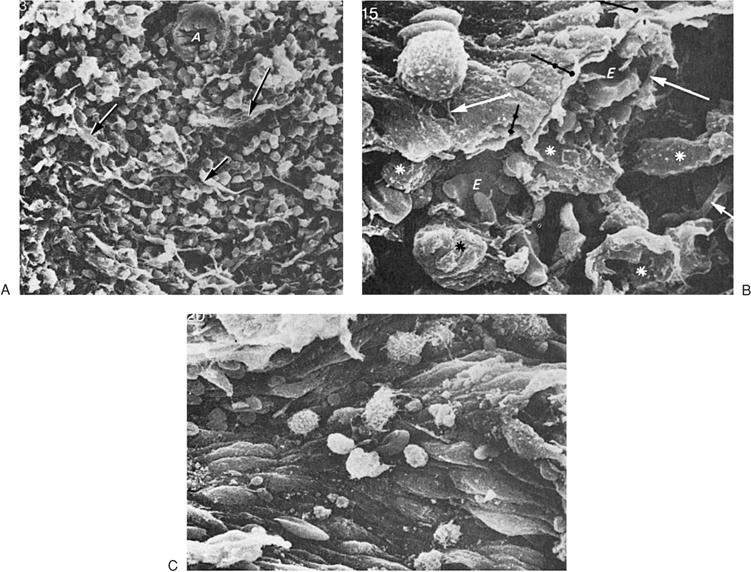
It was suggested originally that human peripheral blood lymphocytes could be separated into two broad categories on the basis of findings of scanning electron microscopy, depending on their cell-surface morphology (20). One population had a fairly smooth surface, whereas the other was described as hairy, being covered by numerous microvilli (Fig. 14.3). The former was considered to correspond to thymus-derived (or T) lymphocyte, whereas the latter was thought to represent bone marrow–derived (or B) lymphocyte. Results of subsequent studies demonstrated that the surface features of lymphocytes depend on the methods used for preparation of the cells as well as the functional state of the cells (21). Thus, lymphocytes stimulated by various mitogens have villi independent of their origin. In addition, both T and B lymphocytes have microvilli while in circulation and especially when they pass through the venules of LNs characterized by high endothelial cells. On the other hand, all lymphocytes have a smooth surface when they reach their respective home microenvironment. Therefore, the smooth cell surface likely is associated with resting lymphocytes, whereas the appearance of microvilli is triggered by environmental stimuli that interact with cell-surface receptors. These newly formed microvilli may help lymphocytes to interact with target substrates.
|
|
|
Figure 14.3. Heterogeneity of human lymphocytes by scanning electron microscope. A: A smooth and a villous lymphocyte, thought to correspond to that of T and B lymphocytes, respectively. The appearance of the lymphocytes, however, depends on the method of preparation; both lymphocytes are smooth when they rest in their microenvironments (×14,500). B: Two villous cells in a preparation depleted of T cells (×14,500). (From Zucker-Franklin D, Greaves MF, Grossi CF. Atlas of blood cells, 2nd ed. Philadelphia: Lea & Febiger, 1988, with permission.) |
![]() Lymphocyte Locomotion
Lymphocyte Locomotion
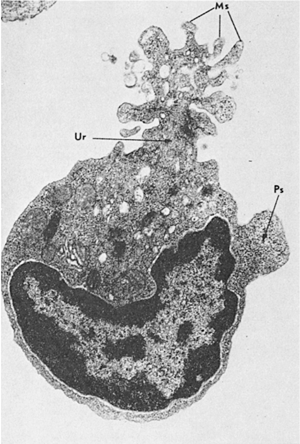
Lymphocyte locomotion was described vividly by Lewis (22), who observed that the cell first extends a pseudopod separated from the rest of the cell by a groove that encircles the cell body. Eventually, as the groove deepens, the nucleus is pushed forward through the constriction ring, giving rise to the classic shape of the motile lymphocyte, resembling a hand mirror or pear. The advancing front is occupied by the nucleus, which is separated by a deep constriction from the rest of the cytoplasm that trails behind, forming the handle of the mirror. The cytoplasmic tail is called the uropod (from Greek ura, “tail,” and podi, “foot”) (Fig. 14.4) (23). It is studded with microvilli and microspikes, which vary in width from 12.5 to 200 nm and to as much as 0.8 μm in length. The motile lymphocyte moves with an average speed of 20 μm/minute. In general, motility increases with the state of cell activation as lymphoblasts demonstrate greater locomotion than small lymphocytes. It could therefore be stated that locomotion is enhanced by stimuli inducing blast transformation. The uropod not only is associated with locomotion, but also serves as a site for a variety of interactions with lymphocytes, macrophages, and other cells (24). In mixed lymphocyte cultures, lymphocytes may approach macrophages and then turn around to establish contacts with them through their uropods. The locomotion of lymphocytes around macrophages has been called peripolesis to distinguish it from emperipolesis (“inside roundabout wandering”), which describes the penetration of malignant cells by lymphocytes. T lymphocytes also adhere to macrophages by means of uropods, forming clusters or rosettes (25). A stable cluster forms only when macrophages and lymphocytes are exposed to antigen. The T lymphocytes within clusters are stimulated into proliferative activity. In general, uropod formation is a morphologic expression of lymphocyte activation as measured by DNA synthesis.
The locomotion of lymphocytes is strikingly different from that of other cells. Lymphocytes move forward in a steady manner, maintaining the hand-mirror shape, whereas myeloblasts express a wriggling wormlike locomotion, and the cells of the monocytic series change shape and direction continuously.
Although it was suggested that uropod formation is a characteristic of T lymphocytes, B lymphocytes also form uropods after stimulation with anti-immunoglobulin antibodies (24). In this case, the uropod forms after capping of the surface immunoglobulin (Ig) is completed; the cap ultimately is found over the uropod, where it is endocytosed. The uropod also has been known to serve as a site for endocytosis of foreign substances. Ultrastructurally, the uropod contains practically all of the cytoplasmic organelles, including the Golgi apparatus, mitochondria, microfilaments, and microtubules.
|
|
|
Figure 14.4. Lymphocyte locomotion. The lymphocyte assumes the characteristic hand-mirror configuration. The nucleus occupies the front, and part of the cytoplasm forms a tail or uropod (Ur), which displays an elaborate pattern of microspikes (Ms). A small pseudopod (Ps) on the side contains only ribosomes. (From Rosenstreich DI, Shevach E, Green I, et al. The uropod-bearing lymphocyte of the guinea pig. Evidence for thymic origin. J Exp Med 1972;135:1037, with permission.) |
|
|
|
Figure 14.5. Plasma cells. A: Normal plasma cell. B: Plasmacytes with vacuoles from the bone marrow of a patient with infection and arthritis. C: Needle type of inclusions in plasma cell. (C used by permission of the American Society of Hematology Slide Bank, 3rd ed., 1990.) |
![]() Plasma Cells
Plasma Cells
Plasma cells are progeny of lymphocytes, although originally they were considered to constitute a separate and independent cell line. Morphologically, they are differentiated easily from other cell types (Fig. 14.5A). The cells are spherical or ellipsoid and range from 5 to 30 μm in size. The cytoplasm is abundant, always is basophilic, and usually is deep blue; it may have a granular character. Plasma cells have a well-defined perinuclear clear zone that contains the Golgi apparatus. In supravitally prepared films, the cytoplasm of plasma cells is deep yellowish gray. The nucleus is small in relation to the cell size; it is round or oval, is eccentrically placed, and contains dense masses of chromatin, often arranged in a wheel-spoke fashion (Radkern).
With use of the electron microscope (Fig. 14.6A), the surface membrane and the nucleus appear to be similar to those of the lymphocyte. The cytoplasm of the plasma cell is characterized by a well-developed rough endoplasmic reticulum that fills most of the cytoplasmic space, except in the area of the perinuclear clear zone containing the Golgi apparatus. The endoplasmic reticulum consists of lamellae arranged in various patterns, usually in parallel convolutions. The inner surfaces of the lamellae are smooth and form the walls of spaces (cisternae) that are filled with amorphous products of varying density. The outer aspects of the lamellae are rough because of attached ribosomes. A few mitochondria may be seen.
|
|
|
Figure 14.6. Electron micrographs of a plasma cell and an intermediate form. A: Typical plasma cell has cytoplasm filled with endoplasmic reticulum (×6,000). B: A lymphocytoid-plasma cell with scant amounts of cytoplasm filled with endoplasmic reticulum and a few mitochondria (m) (×9,900). N, nucleus. (From Maldonado JE, Kyle RA, Brown AL, et al. “Intermediate” cell types and mixed cell proliferation in multiple myeloma: electron microscopic observations. Blood 1966;27:212–226, with permission.) |
The above-described electron microscopic appearance of plasma cells is the most typical. Often, however, intermediate forms are found (14). Thus, some cells may resemble small lymphocytes morphologically but may contain an unusually well-developed rough endoplasmic reticulum (Fig. 14.6B). Such intermediate cells (Fig. 14.1C-E) are common in the blood of patients with plasma cell dyscrasias and of those with immunologic diseases characterized by hypergammaglobulinemia (27). Similar cells have been encountered in the blood of patients with viral infections (Turk cells) (19), including infectious mononucleosis, as well as in the blood of apparently healthy individuals. Alternatively, immature plasma cells may have an appearance more akin to that of phytohemagglutinin (PHA)-transformed cells; the nucleus is large and leptochromatic and the cytoplasm contains a rather simple endoplasmic reticulum, but many ribosomes and polyribosomes are present. Thus, it is often difficult to draw sharp cytologic dividing lines. Other plasma cells may contain vacuoles or needle-type inclusions in the cytoplasm (Fig. 14.5B,C). The molecular and genomic aspects of plasma cell differentiation are discussed in Chapter 16.
![]() Lymphoid Organs
Lymphoid Organs
The tissue aggregates of the immune system are separated into primary and secondary lymphoid organs. In humans, the primary lymphoid organs are the bone marrow and the thymus. The secondary lymphoid organs are the spleen, the LNs, the Peyer patches (PPs) of the gut, and the Waldeyer ring (tonsils and adenoids). This division provides an anatomic basis for two fundamental stages of lymphocyte differentiation: The antigen-independent stage and the antigen-dependent stage. The primary lymphoid organs develop before the secondary organs in ontogeny and provide the proper microenvironment for antigen-independent differentiation of lymphocytes from immature precursors. At the end of this early stage of lymphocyte differentiation, immunocompetent lymphocytes are released and home to specific areas of the secondary lymphoid organs. The secondary lymphoid organs provide an optimal microenvironment for attracting antigen-specific lymphocytes, directing the terminal stages of lymphocyte differentiation, and distributing the fully differentiated effector cells or their products to other parts of the body.
Primary Lymphoid Organs
Bone Marrow
A detailed account of the histologic features and function of bone marrow is given in Chapter 5. Here we concentrate on the contribution of bone marrow in lymphopoiesis as a “primary lymphoid organ.” The bone marrow occupies the medullary cavities of bones throughout the skeleton. It is the major hematopoietic organ in humans and supports differentiation of all blood cells (28). In some cases, such differentiation is not always complete. T lymphocytes and monocytes, for example, reach their final stages of maturation in locations outside the bone marrow. The bone marrow is divided into an extravascular compartment, which is the site of hematopoiesis, and a vascular compartment, composed of wide venous blood vessels known as sinuses. These vessels receive blood from the nutrient artery and the periosteal capillary network. The sinuses are radially disposed in the bone marrow and eventually open into larger, centrally located sinuses that exit through the same foramina used by the nutrient arteries. The walls of the venous sinuses consist of an endothelial layer, a basement membrane, and the adventitia. The endothelial cells are flat, with tapering ends, and contain the usual organelles. They contain lysosomes and therefore are endowed with endocytic activity. The adventitial cells have broad sheetlike processes that form a reticulum, the interstices of which are occupied by the hematopoietic cells. Under some circumstances, the adventitial cells become swollen because of an increased fat content, and the gross appearance of the marrow turns from red to yellow. In conditions of increased demand on the hematopoietic process, the fatty adventitial cells decrease in size, allowing an expansion of the hematopoietic compartment. The adventitial cells are related to the reticular cells found in the splenic cords and are therefore called adventitial reticular cells.
The hematopoietic compartment displays a specific cellular arrangement of hematopoietic cells. For example, the megakaryocytes lie close to the adventitial cells and deliver platelets directly into the sinuses through apertures in the sinus walls. Erythrocytes are produced near the sinuses, forming erythroblastic islets. Granulocytopoiesis takes place at a distance from the sinus wall in a diffuse pattern or sometimes in distinct clusters or sheets. Available evidence obtained through labeling techniques suggests that new lymphocytes are formed at the periphery of the bone marrow and move toward the center in a centripetal fashion. The lymphocyte progenitors are enriched in a prolymphocyte population Lin-c-kitlo that contains progenitors for B cells, which are CD122- (interleukin [IL]-2β), and for natural killer (NK) cells, which are CD122+, and under defined culture conditions can mature toward B or NK lineage. B cells are generated from hematopoietic stem cells (HSCs), which migrate from the subendosteal region toward the center of the cavity. These stem cells differentiate toward several cell lines as a result of signals received from bone marrow–adherent cells that create microenvironments known as niches that support hematopoiesis. The bone marrow stromal cells can be identified by their phenotype (i.e., CD105, CD90, CD184 and class II major histocompatibility complex [MHC] antigens) (29). HSCs apparently interact with the osteoblasts of the bone, which may contribute to the creation of the hematopoietic niche (30). Osteoblasts may also support the development of hematopoietic colonies as a result of secretion of a variety of cytokines (31). The development of an in vitro culture system by Whitlock and Witte (32) facilitated the identification of the growth and differentiation factors necessary for B-cell lymphopoiesis. Animal studies defined five stages of B-cell differentiation downstream from the common lymphocyte progenitor (CLP) cell: (a) prepro–B cell, (b) pro–B cell, (c) pre–B cell, (d) immature B cell, and (e) mature B cell. The CLP cell is the first one irreversibly committed to B- or T-cell differentiation. What drives the terminal stages of B-cell differentiation within certain bone marrow niches is the availability of certain growth and differentiation factors such as Ikaros (33), early B-cell factor (EBF), and PAX-5 (34,35). The FLT-3 ligand is essential for the generation of CLPs and prepro–B and pro–B stages, while the chemokine ligand CXCL-12 (also known as stromal cell–derived factor-1 [SDF-1]) is essential for prepro–B- to pro–B-cell stages (36). Interleukin-7 (IL-7) contributes to the growth of stages after pro–B cell. The chemokine CXCL-12 was isolated from stromal cell lines used for studying the B-cell development in vitro and with its receptor CXCR4 is essential for B-cell development and other developmental processes such as angiogenesis and neurogenesis. The CXCR4 receptor functions also in guiding and retaining B cells to appropriate secondary lymphoid organs (37). Mice deficient in IL-7 or the IL-7Rα have severe deficiency of B (and T) cells. IL-7 is required especially at the pro–B-cell stage (38). The labeling index of lymphocytes within the sinuses is intermediate between those of the whole bone marrow and those in the peripheral blood, suggesting that newly synthesized lymphocytes are discharged into the circulation. Identifiable lymphocytes are found singly or in small groups near the sinusoidal walls, with some of them in transit through the wall of the sinus. B lymphocytes are not stored in the bone marrow except for brief periods before their release into the circulation. Histologically, no lymphoid follicles are distinguishable in the normal bone marrow.
The bone marrow resembles the red pulp of the spleen with its vascular sinuses and a reticular meshwork formed by the adventitial cells or the cordal reticular cells, respectively. It differs in that the bone marrow circulation is closed; the circulation of the red pulp is at least partially open, with no endothelial continuity between the arteries and the veins.
Thymus
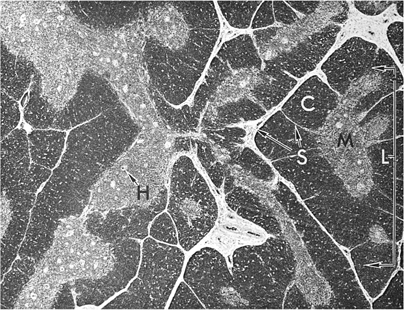
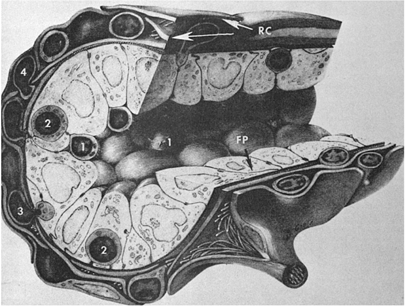
The thymus is a lymphoepithelial organ situated in the superior mediastinum (39,40). It consists of two lobes that are divided into lobules that form the basic anatomic units of the thymus. The thymus is covered by a fibrous capsule from which fibrous bands (trabeculae) penetrate the parenchyma, dividing it into lobules. Histologically, the lobules have two distinct regions (Fig. 14.7): The peripheral region, which is called the cortex and is divided into the outermost or subcapsular cortex and the inner or deep cortex, and the central region, which is called the medulla. In hematoxylin-eosin–stained sections, the cortex appears dark blue to purple because of the predominance of lymphocytes (80 to 85%), whereas the medulla appears eosinophilic because of the predominance of the epithelial cells. These anatomic divisions correspond to functionally distinct microenvironments that support specific phases of thymocyte differentiation. The subcapsular or superficial cortex contains large, actively dividing blasts with many mitotic figures. The deep cortex is composed principally of nondividing, small thymocytes, and the medulla contains predominantly medium-sized thymocytes. The relationships between the thymocytes in each compartment and the mechanisms underlying thymocyte differentiation are discussed in detail in Chapter 16.
|
|
|
Figure 14.7. Histology of thymus shows lobules (L) consisting of cortex (C) and medulla (M). H, Hassall corpuscles; S, septa (trabeculae). |
Ontogeny
The embryologic origins of the thymus have been under dispute for more than 30 years (41). According to one view, the thymic primordium arises from the third pharyngeal pouch (endoderm) as well as the pharyngeal crest (ectoderm) (42), while another study supported exclusively the endodermal origin (43).
More recently, endodermal labeling with cell tracker dye demonstrated that endoderm alone contributes to the formation of the thymus (44). The pharyngeal pouches are transient embryonic structures and their formation requires the paired box gene 1 (Pax 1) and 9 (Pax 9) as well as the fibroblastic growth factor 8 (Fgf8). They develop in an anterior to posterior sequence and are separated from each other by pharyngeal arches. The pouches contribute to the formation of several organs including the thymus and the parathyroids. Each endodermal primordium contains precursors for one thymus lobe and one parathyroid gland. It is surrounded by a capsule of mesenchymal cells derived from cells of the neural crest, which supports the growth and development of the primordium, and under their guidance it separates into a thymus and pharynx. The development of the thymus proceeds through several stages: (a) positioning (i.e., the determination of the place where the organ rudiment will develop); (b) growth and patterning (i.e., generation of distinct regions in the growing rudiment); and (c) differentiation (i.e., separation of distinct cell types that have been developed). The function of the homeobox A3 (HoxA3) is necessary to identify the third pouch, which forms the thymus and may control the position of the mitral rudiment (44). Mice lacking HoxA3 are athymic and they have no parathyroids since the third pouch endoderm cannot differentiate appropriately.
In all pharyngeal pouches Pax1 expression requires two other genes such as the Eyes Absent gene (Eya1) and the sine oculus-related homeobox1 homologue (Six1) (45). Functional evidence that the endoderm alone forms fully functional thymus was obtained through ectopic transplantation of isolated endoderm tissue grafted under the kidney capsule (46).
Normally, the original tubular epithelial structures, one on each side, descend into the anterior superior mediastinum where they fuse, forming the mature organ with two lobes. At this stage, the thymus is triangular, with the base resting on the pericardium and the apex pointing toward the neck. Once the anlage is formed, the function of the whn (winged helix nude) gene is required (47,48). The whn gene encodes a nuclear protein, Whn, which is a transcription factor of the forked/winged/helix family. It contains a DNA binding domain and a strongly acidic transcriptional activation domain, which is functionally indispensable. Mutations affect the DNA binding domain or the C-terminal transcriptional activation domain and are associated with athymia and hairlessness because the gene regulates the keratinization of hair.
Mesenchyme of neural crest origin also plays an important role in thymic development (49). It migrates into the thymus early in development, surrounding the epithelial rudiments, and is closely associated with the developing thymocytes. It forms the connective tissue capsule and trabecular septa that penetrate the parenchyma, dividing it into lobules (49). It is also likely that it provides extracellular matrix into the cortex and medulla, such as hyaluran, collagen, and fibronectin, which may be important for the presentation of growth factors to the thymocytes (50). This part of mesenchymal development occurs after the lymphocytic invasion.
The undifferentiated epithelial cells are large, with dispersed chromatin; short, blunt cytoplasmic processes; and sparse tonofilaments. As the cell matures, the amount of cytoplasm decreases, and long dendritic processes form, which permit tight junctions between adjacent cells and the formation of the epithelial framework within the interstices of which T-lymphocyte differentiation takes place.
Migrating lymphoid progenitors initiate a symbiotic relationship with the epithelial cells and contribute to the organization of the thymic microenvironments. Whereas prothymocytes (i.e., immature cells) regulate induction of the cortical microenvironment (51), mature T-cell receptor (TCR)+ thymocytes organize thymic medullary epithelial cells (52).
E-cadherin plays an important role at this stage because it mediates the interactions that shape the architecturally distinct cortical and medullary thymic microenvironments (53). Heterotypic interactions occur between epithelial cells expressing E-cadherin and double-negative T lymphocytes expressing the ligand of E-cadherin (i.e., CD103 [αEβ7 integrin]). These interactions enhance thymocyte proliferation (54,55).
The thymic anlage is permeated by blood vessels, which also contribute to a well-organized medullary epithelial compartment, even in the absence of CD3+ mature thymocytes (i.e., in Rag-2-/- mice). These animals develop a hypoplastic thymus (due to lack of mature T cells), but the medullary epithelium is otherwise well organized. Addition of keratinocyte growth factor, a member of the fibroblast growth factor family secreted from mesenchymal cells, results in the expansion of the epithelial medullary compartment in the Rag2-/- mice (56). The relB gene encoding a subunit of the nuclear factor-κB complex is critical in coordinating several aspects of the development and organization of medullary epithelial cells and dendritic cells (DCs).
Overexpression of CD40L on thymocytes disrupts the thymic architecture and the epithelial differentiation, with loss of cortical epithelial cells, expansion of the medullary compartment, and infiltration of the capsule with a mixture of CD3+ cells, B cells, and macrophages (57). These findings point out that regulation of normal development of thymus is a highly complex process, delicately regulated by multiple cellular interactions and soluble factors released as a result of these interactions.
The medullary epithelium expresses several molecules, which are considered tissue specific, such as parathyroid hormone, thyroglobulin, insulin, and C-reactive protein, and even structures with features of respiratory epithelium and others, which phenotypically and structurally resemble thyroid follicles. This may represent a “promiscuous” gene expression or a “mosaic” of epithelial differentiation (58), which may form the basis for the establishment of self-tolerance. This view is supported by the demonstration that the gene aire, which is defective in autoimmune polyendocrinopathy candidiasis–ectodermal dystrophy (APECED), is expressed by medullary thymic epithelial cells (59).
AIRE (auto immune regulator) is a DNA-binding nuclear protein with a putative DNA-binding domain named SAND (Sp100, Aire-1, NucP41/75, DEAF-1) and four, LXXLL nuclear receptor binding motifs (shown here in amino acid one-letter code, with X denoting any amino acid), a structure indicating that AIRE functions as a transcription regulator (60,61).
It interacts in vitro with CREB-binding protein (CBP), a property that mediates its transcriptional regulation (62). It also contains a proline-rich region, known to be expressed by several transcription factors, mediating interaction with SH3 domains.
The LXXLL motif binds coactivators to nuclear receptors, thus activating transcription of target genes by nuclear receptors. AIRE plays a vital role in preventing autoimmunity, and AIRE mutations cause a recessive inherited disease known as APECED or autoimmune polyendocrinopathy syndrome type 1 (APS-1). APECED is expressed at the beginning as mucocutaneous candidiasis early in childhood with eventual destruction of endocrine organs and the appearance of Addison disease, hypoparathyroidism, and type I diabetes (63). AIRE is expressed in medullary epithelial cells, testes, and cells of dendritic lineage (64). It has been suggested that AIRE may induce the transcription of genes specific to other tissues that are promiscuously expressed in the thymic epithelial cells (65) (see Chapter 16). It has been suggested that AIRE promotes the transcription and expression by the thymic epithelial cells of self-antigens from other tissues facilitating the removal of thymocytes with self-reactive T-cell receptors (66,67). The thymus is located within the upper part of the thoracic cavity. Recently in mice a second thymus has been identified in the neck with typical cortical and medullary structures (68). The cervical thymus is structurally the same as the thoracic thymus; that is, it consists of cortex and medulla and contains double-positive (CD4+/CD8+) and single-positive (CD4+/CD8- and CD4-/CD8+) thymocytes, expressing TCR/CD3 complexes. The cervical thymus is fully functional because it could supply T cells to athymic (nude) mice, correcting their immune deficiency. This unexpected discovery poses a number of questions on the vast amount of experimental work done with thymectomized animals over the past 45 years (69). Perhaps the cervical thymus is too small to be relevant and its cellularity is only 1/500th of the thoracic thymus (70). However some of the work done with thymectomized animals, such as the role of the thymus in autoimmune disease, may have been reconsidered (71).
Nonlymphoid Cellular Elements
Epithelial Cells
The thymic rudiment is populated by epithelial cells that anatomically are distinguished as cortical and medullary. As indicated earlier, lack of Hoxa3 and Eya1 genes impact the early phases of thymic organogenesis from the third pharyngeal pouch. The epithelial compartment of the developing thymus is regulated by Pax1 (72), Pax9 (73), and Foxn1 (74) genes. Deletion of Pax1 results in thymic hypoplasia as a result of defective organogenesis of the thymus, while Pax9 causes marked thymic hypoplasia because of defective cellular migration to the thymic rudiment. Loss of Foxn1 function, on the other hand, results in a more complex phenotype known collectively as the “nude” mouse. The “nude” mouse has been an important “experiment” of nature that has provided important insights on the function of thymus to generations of immunologists. The abnormal thymic development (75) is associated with defective keratinization (76), and although the early stages in thymic development proceed normally, including normal mediastinal migration, the subsequent thymic rudiment does not develop, as a result of failure of its colonization by lymphoid progenitors (77).
It is generally accepted that thymic organogenesis is completed in two distinct steps. During the first stage, the thymic rudiment is formed from endodermal elements of the third pharyngeal pouch and migrates to the mediastinum. In the second stage the Foxn1 transcription factor drives thymic cell lineage differentiation from multipotential progenitors. In the absence of Foxn1 function, the cells of the thymic rudiment phenotypically and in their organizational features resemble respiratory epithelium (74).
|
|
|
Figure 14.8. Ultrastructure of the thymus. An epithelial dendritic cell at center (E) extends long cytoplasmic processes (arrows). Note the intimate contact between the epithelial cell and the surrounding thymocytes (×4,400). (From Bloodworth JMB Jr, et al. Ultrastructure of the human thymus, thymic tumors, and myasthenia gravis. Pathol Annu 1975;10:329, with permission.) |
Expression of Foxn1 is regulated by products of the Wnt genes, which play a key role in the regulation of cell fate, specification, proliferation, migration, and death (78). The Wnt glycoproteins bind to glycosaminoglycans of the extracellular matrix, cell-surface receptors of the Frizzled (Fz) family, and members of receptors related to low-density-lipoprotein receptors.
Through the regulation of Foxn1 function the Wnt proteins contribute importantly in the development of thymic epithelial cells. The epithelial cells in the cortex and medulla, although morphologically distinct, have a common progenitor as shown by clonal analysis (79), and such progenitors still exist in the thymus after birth (80).
Ultrastructurally, the epithelial cells are phenotypically highly heterogeneous, probably reflecting their divergent origins (see Chapter 16). In the past, the epithelial cells have been referred to by a variety of designations, such as reticular, syncytial, epithelioid, and squamoid (Fig. 14.8). The characteristic features that distinguish them from other cells in the thymic parenchyma are the presence of tonofilaments and desmosomes. The tonofilaments are filamentous structures approximately 3.0 nm in diameter and 0.1 to 3.0 μm in length. They often form bundles and are located close to the nucleus. Desmosomes are detected at the junctions of the dendritic processes of adjacent epithelial cells. Their oval nuclei have evenly distributed chromatin and prominent nucleoli. The epithelial cells joined by desmosomes form a continuous meshwork that surrounds the trabeculae, the blood vessels, and the inner surface of the capsule. Between the epithelial sheath and the supporting scaffolding of the trabeculae and blood vessels is a continuous basement membrane distinct from the basement membrane of the capillaries. The epithelial meshwork thus separates cavernous spaces filled with thymocytes.
The cortical epithelial cells do not have secretory granules and probably do not participate in the secretion of thymic hormones. Such interactions are regulated by products of genes of the major histocompatibility complex. Indeed, the presence on epithelial cells of class I and II molecules in mice and humans has been demonstrated. The distribution of class II antigens is typically dendritic in the cortex, but it is confluent in the medulla (81). The class II antigens are of higher density than the class I antigens in cortical epithelial cells, but they are of the same high density in the medullary epithelium. Using immunohistochemical techniques and electron microscopy, investigators have shown that these molecules are of epithelial origin because they are located on cells containing desmosomes and tonofilaments.
|
|
|
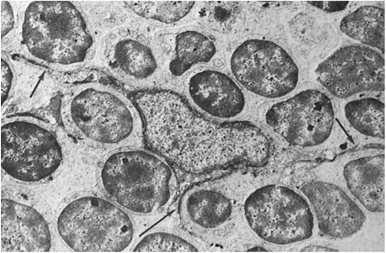
Figure 14.9. A typical large nurse cell containing many thymocytes (L) and two epithelial nuclei (N). Between the internalized thymocytes are cytoplasmic vesicles and tonofilaments. Some nurse cells contain thymocytes in mitosis. (From Wekerle H, Ketelsen UP, Ernst M. Thymic nurse cells. Lymphoepithelial cell complexes in murine thymuses: morphological and serological characterization. J Exp Med 1980;151:925, with permission.) |
The thymocyte–epithelial cell interaction in the outer cortex results in the formation of lymphoepithelial complexes known as nurse cells (Fig. 14.9) (82). They are composed of large clusters of lymphocytes surrounded by cell membrane and appear to reside within the cell body of an epithelial cell. Their significance in T-cell differentiation is discussed in Chapter 16. The epithelial cells of the medulla do not have long dendritic processes and are known as epithelioid or spatulate epithelial cells. They are more pleomorphic and contain dark granular inclusions that may have secretory functions. Another variety of epithelial cells, known as squamoid, is found exclusively in the medulla. These cells probably give rise to Hassall corpuscles and contain dense bundles of tonofilaments and masses of keratohyalin.
The Hassall corpuscles described first more than 150 years ago (83) are solid or cystic, and their origin is highly disputed. In 1906, Hammar suggested that Hassall corpuscles arise as solid structures through the tight apposition of as many as 20 to 50 epithelial cells in a concentric fashion. Degenerative changes in the centrally located cells produce the cystic forms. The cavity of the cystic forms of these corpuscles is lined with epithelial cells with villous projections, and sometimes it is filled with debris and degenerated cells. In some solid corpuscles, keratinization is prominent and is similar to that of the skin. Monoclonal antibodies specific for high-molecular-weight keratins that react with terminally differentiated epithelial cells of the skin also bind to Hassall corpuscles (39,84). Epithelial cells of the deep cortex are phenotypically different from those in the medulla, which undergo terminal keratinocyte differentiation. As shown recently, Hassall corpuscles are not the epithelial cell graveyard as previously thought, but contribute actively to important immunoregulatory functions. They secrete thymic stromal lymphopoietin (TSLP), which regulates CD4+ T-cell homeostasis through activation of thymic dendritic cells (TDCs) (see below). It has now been shown that TSLP induces proliferation and differentiation of CD4+/CD25+/Foxp3+ regulatory T cells from CD4+/CD25- thymocytes (85).
Macrophages
Macrophages are found throughout the thymus: Among thymocytes, in capsular and septal connective tissues, and in the perivascular space. Some macrophages are surrounded by thymocytes and form rosettelike structures. Thymic macrophages direct thymocyte maturation, but appear to be involved also in the disposal of many thymocytes dying within the thymus. Macrophages are more prominent in involuted thymuses after stress or steroid hormone treatment, rendering the histologic picture of a starry sky. Lymphocyte phagocytosis is found in the germinal center (GC), a site that also is characterized by high proliferative activity.
Interdigitating Dendritic Cells
A third population of nonlymphoid cells in the corticomedullary junction is known as interdigitating dendritic cells or thymic dendritic cells (TDCs). They contain an irregularly shaped nucleus and clear cytoplasm, but they lack the Birbeck granules characteristic of Langerhans cells (LCs), and their phenotype and origin apparently varies. Some are CD8a+, derived from a common precursor with T cells (86), and therefore are lymphoid related; that is, granulocyte-macrophage colony-stimulating factor is not required for their development (in contrast to the myeloid DCs), and they need functional Ikaros transcription factors for their maturation (87).
In humans, pluripotent stem cells, which leave the bone marrow to migrate to the thymus, have the potential to develop into T cells and DCs (88).
Other TDCs express the DC116+/CD8α- phenotype (i.e., that of Langerhans cells in other tissues) and originate from DC precursors (89). Their differentiation is driven by factors produced by thymic epithelial cells as well as Hassall corpuscles (90). Another pathway of TDC differentiation is from double-negative thymocytes (i.e., CD4-/CD8-) stimulated, in the presence of IL-7, by IL-18 produced by thymic epithelial cells (91).
Thymic stromal cells produce the IL-7–like lymphopoietin TSLP that acts through appropriate receptors on dendritic cells and induces expansion of CD4+ T cells carrying self-peptide–MHC complexes (92). TSLP is also produced by epithelial cells in tonsillar crypts and other mucosal tissues, and thus may exert a homeostatic mechanism on CD4+ T cells, through dendritic cell activation not only within the thymus, but in other mucosal lymphoid tissues as well.
Myoid Cells
Myoid cells contain numerous intracytoplasmic filaments arranged in a haphazard fashion and are seen in many lower vertebrates. Their presence in human fetal thymuses has been documented, but their identification in postnatal thymuses remains controversial. One suggestion is that myoid cells originate from mesenchymal or epithelial cells. The latter seems likely because of their attachment to epithelial cells by desmosomes.
Lymphoid Cells
Lymphoid cells constitute 80 to 85% of the cortical and 15% of the medullary cells. The outer cortex lymphocytes are large blast cells with dark blue cytoplasm that proliferate actively. They are the immediate descendants of bone marrow–derived prothymocytes, which enter the thymus and migrate to the subcapsular cortex. As they mature, they move into the deeper cortex, which is occupied by small nondividing thymocytes. The medullary thymocytes are of medium size and are considered emigrant cells, although this fact has not been proven conclusively. The recruitment, migration, and differentiation of the lymphocytes and the relationship between subcapsular, deep cortical, and medullary thymocytes are discussed in Chapter 16.
Thymic Vasculature: Blood–Thymus Barrier
The thymic anlage initially is avascular. Vessels penetrate the thymus at approximately 12 to 14 weeks of gestation. The thymic arteries, which are branches of the inferior thyroid, internal mammary, and pericardial phrenic arteries, enter the thymus from the surrounding tissues, pass down to the medulla, and branch into arterioles that penetrate into the deep cortex. Capillaries that arise from the arterioles run toward the subcapsular cortex, where they anastomose and turn inward toward the medulla, eventually forming venules (93). The vessels are ensheathed with epithelial cells and connective tissue that is continuous with the capsule. Histologically, therefore, a number of layers can be distinguished from the lumen of the blood vessels outward: Endothelium; the vascular basement membrane; a mesenchymal perivascular connective tissue space occupied by collagen fibers, fibroblasts, some macrophages, and other cells; an epithelial basement membrane; and the epithelial cell syncytium. These layers constitute the blood–thymus barrier. The mesenchymal interstitial space is probably formed in the early stages of development when the thymic epithelial cells envelop the blood vessels. This space is considered extraparenchymal. The tightness of the barrier has been tested with particulate and protein tracers. It is tight in the cortex, mainly because of the impermeability of the endothelial junctions. Traces of protein transported by plasmalemmal vesicles of the endothelial cells and released on the parenchymal front are removed rapidly by the macrophages along the vessels in the interstitial space and thereby are prevented from coming into contact with the thymocytes. The barrier is incomplete in the medulla, however, especially along the site of thymocyte migration through the wall of venules (94). The fact that antigens can be detected in the thymus indicates that the “barrier” has leaks (95). Nevertheless, experimental tracers never reach the cortical parenchyma. This arrangement of the vasculature and epithelial sheaths separates the thymus into the intraparenchymal compartment, composed of the lymphoepithelial complex, and the extraparenchymal compartment, composed of the blood vessels and the surrounding interstitial space.
Involution
Beginning at puberty, the thymus undergoes a gradual process of involution, characterized by loss of the cortical lymphocytes and atrophy of epithelial cells and their replacement by fat originating from mesenchymal cells present in the connective tissues along the vasculature and capsule. More than 50% of the thymus is replaced by adipose tissue by the age of 40 to 45 years, but the fat is still contained in the extraparenchymal compartment, separated from the remaining lymphoepithelial complex by the epithelial basement membrane and the sheet of epithelial cells. The speed of age-related involution may be influenced by stress and other factors.
Thymic involution starting at puberty has been associated with use of sex hormones; however, this hypothesis has been challenged since morphometric studies have shown that thymic involution actually starts from the first few years of life (96). It is a continuous process affected by the decrease of the potential of intrathymic progenitors to maintain the level of mature thymocytes (97).
Associated with rise of sex hormone concentration is a decline in the production of growth hormone, and many of its effects in the periphery are mediated by insulin growth factor-I induction (98). Thymopoiesis is maintained throughout life with complete, although slow, renewal of T cells (99). Therefore, changes of the thymic tissue are quantitative rather than qualitative. Thymic involution is linked at the molecular level with expression of JAGGED-1 in T cells (100). JAGGED and Delta proteins are members of the Delta/Serrate/Lag2 (DSL) genes that regulate Notch signaling. Notch signaling is implicated in regulation of T-cell activation and differentiation. JAGGED-1 expression on T cells interferes with Notch signaling and induces premature involution as a result of thymic epithelial cell apoptosis.
In contrast to the chronic involution occurring as a result of aging, the thymus undergoes acute involution as a result of stress. This involution is mediated by adrenal corticosteroids. Injections of glucocorticoids eliminate as much as 75% of the thymocytes within 2 to 3 days, affecting both the cortex and the medulla, but the effects are more pronounced in the cortex. Most of the cortical thymocytes are cortisone sensitive, whereas the medullary thymocytes are cortisone resistant. After acute stress, the lymphocytes undergo pyknotic changes, fragmentation of the nuclei, and a decrease in size; eventually, they are phagocytosed by macrophages. Recovery takes place within 8 to 10 days and is marked by an early increase in mitotic activity in the subcapsular cortex.
Pathology of the Human Thymus
Myasthenia Gravis
The thymus from patients with myasthenia gravis typically shows histologic changes characterized by the development of follicles with GCs composed of B cells typical of reactive peripheral lymphoid organs (101). They are located outside the epithelial basement membrane, predominantly in the medulla, and are surrounded by many Hassall corpuscles. Many plasma cells are easily discernible. In some cases, an increase in the number of medullary epithelial cells containing thymic hormones is noted. The cause of these histopathologic lesions is unknown, but antibodies against the acetylcholine receptor in the neuromuscular junction play an important role in the pathogenesis of myasthenia gravis, and it is intriguing that the receptor, or a structurally similar antigen, has been detected on thymic cells (102). Antibodies from patients with myasthenia gravis react with striations of myoid cells from animal and human thymuses in culture and muscle cells expressing acetylcholine receptors (103). Approximately 10 to 15% of patients with myasthenia gravis have thymomas, but these histologic changes are not characteristic of myasthenia gravis in that they have been observed in patients with other diseases, such as endocrinopathies (Addison disease and thyrotoxicosis) and autoimmune diseases (such as systemic lupus erythematosus).
Similar lesions have been identified in thymuses from patients with multiple sclerosis. A well-documented fact is that the thymus and the brain share some antigens, such as the Thy-1 antigen (CD90).
Dysplasia
The thymus of patients with severe combined immunodeficiency disease is characterized by scant lymphocytes with reduced numbers of epithelial cells.
Thymomas
Tumors of the thymus are epithelial or lymphocytic, or they contain a mixture of both cellular elements. In some cases, ductal and glandular structures are detected, mixed with other epithelial cells. These cases are thought to imitate the early ontogenetic stages of the thymic anlage and support the view of the double origin of the thymic epithelium. Thymic lymphomas that appear as mediastinal masses are associated with T-cell acute lymphatic leukemia in children. The cells of these tumors bear the phenotype of cortical thymocytes in the early stages of their development. The diagnosis of primary thymic epithelial tumors has been controversial, because of the difficulties in their histopathologic classification and prognostication of their clinical behavior. A new histologic classification has recently been proposed by the World Health Organization (WHO) (104), which provides an improved platform, over pre-existing classifications, in clinical practice for assessment and treatment of patients.
Secondary Lymphoid Organs
Lymph Nodes
Ontogeny
The ontogeny of the secondary lymphoid organs, LNs, PPs, and spleen, is highly complex (105,106,107). The fundamental concepts of lymphoid organ neogenesis were generated approximately 100 years ago (108). The first stage in the development of the LN and PP is the budding from large vessels of lymph sacs, which are then penetrated by connective tissue and lymphatic vessels. These vessels interconnect with other vessels, forming a network. The Prox-1 gene is expressed on lymphatic endothelial cells and is required for budding and sprouting of the lymphatic endothelium (109). Sabin (108) observed that one spot of budding to generate a lymph sac occurs in the anterior cardinal veins of the neck. This lymph sac gives rise to the lymphatics of the neck, thorax, heart, lungs, and forelegs. In the same location, the expression of the Prox-1 gene was identified. After the development of the lymphatics, circulating CD45+/CD4+/CD3- hematopoietic, progenitor cells from liver provide the signals for induction of LNs and PPs. Lymphotoxin (LT) is essential for the formation of LNs and PPs, as shown in LTα-/- mice in which there is a profound defect in the formation of LN, complete absence of PP, and grossly disturbed splenic microarchitecture. The membrane form of LT is the active signaling molecule in LN and PP biogenesis (110). LTα forms heterotrimers with LTβ (LTα1β2) and exists as a membrane-bound form on the cell surface. LTα1β2does not bind to either of the two tumor necrosis factor (TNF) receptors but to its own receptor, the LTβ receptor (LTβR) (111). LTα1β2 binds to its own receptor, LTβR, and results in the induction of the nuclear factor–κB (NF-κB)inducing kinase (NIK), initiating an alternative pathway for activation of the transcription factor NF-κB, which plays a pivotal role in innate and adaptive immunity (112). This alternative pathway (i.e., LTα1β2 → LTβR → NIK → inhibitory κ-B kinase) (IKKα) constitutes a major pathway in the development, organization, and function of the secondary lymphoid organs. The chemokines CCL19, CCL21 (which bind to chemokine receptor CCR7), and CXCL13 (which binds to receptor CXCR5) are also important for the colonization of lymphoid organs by T and B cells and antigen-presenting cells (113).
After completion of the connective tissue structure, the first cells to colonize the LN in mice express the phenotype CD4+/CD45+ (114). These cells are not T cells because they are TCR- and CD3- but express LTα1β2. They do not differentiate to either T or B lymphocytes but after stimulation by IL-2, they differentiate to NK cells and with a combination of other cytokines give rise to antigen-presenting cells (115). The precursors of these cells are IL-7R positive and reside in the liver (116). The generation of the CD45+/CD4+/CD3- cells from precursors in the liver requires the function of the RORγ gene (117) as well as the Id2 gene, which in mice regulates LN and PP development (118).
Finally, the development of all LNs (but not PPs) depends on the TRANCE/TRANCE-R signaling. TRANCE (or RANK-L, OPG-L) is a member of the TNF family of cytokines, and mice deficient in TRANCE have absent LNs and a serious defect in bone development due to the absence of osteoclasts (119), but have normal PPs (120).
Mutation of another gene “aly” (alymphoplasia) results in complete absence of LNs and PPs and lack of well-defined lymphoid follicles in the spleen (121).
Development of B-cell–rich lymphoid follicles requires the function of the B-lymphocyte chemoattractant (BLC) or CXCL13 and its receptor CXCR5, which stimulates homing of B lymphocytes to the follicles. BLC induces up-regulation of α1β2 LT, which promotes follicular DC development and BLC expression, establishing a positive feedback loop (122). Colonization of the paracortical areas by T cells is regulated by another chemokine system (see Lymphocyte Homing and Recirculation).
Structure
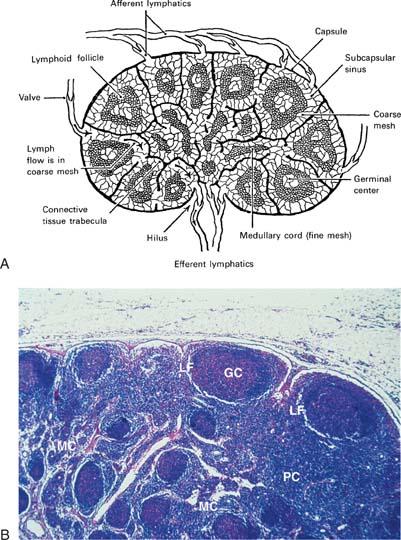
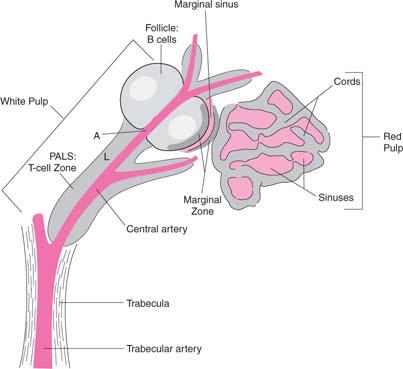
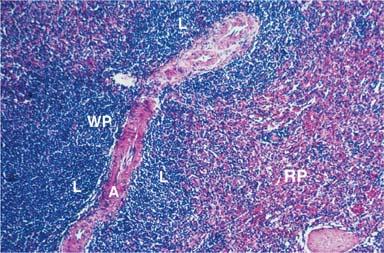
The LNs are ovoid structures ranging in size from a few millimeters to more than a centimeter. They are particularly common at the base of extremities, in the retroperitoneum and the mediastinum, and along blood vessels. LNs are surrounded by a fibrous capsule from which trabeculae penetrate the parenchyma, forming a fibrous supporting meshwork. Blood vessels enter and leave LNs through the hilum. Histologically, two regions can be distinguished: A peripheral cortex and a central medulla. The cortex is subdivided into the superficial cortex, located immediately beneath the capsule, and the deep cortex or paracortex, which is located toward the center of the node (Fig. 14.10). To gain a clear understanding of LN function, it is useful to regard LNs as consisting of the reticulum and the cellular compartment.
|
|
|
Figure 14.10. Lymph node. A: Drawing of a lymph node. B: Cross section of a normal lymph node. GC, germinal centers; LF, lymphocytic follicles; MC, medullary cords; PC, paracortical lymphoid areas. |
Lymph Node Reticulum
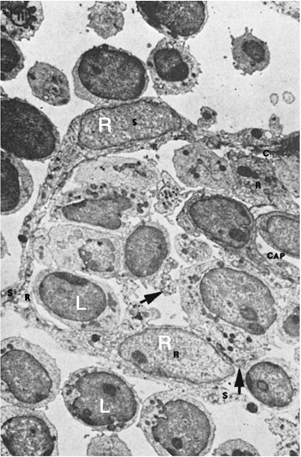
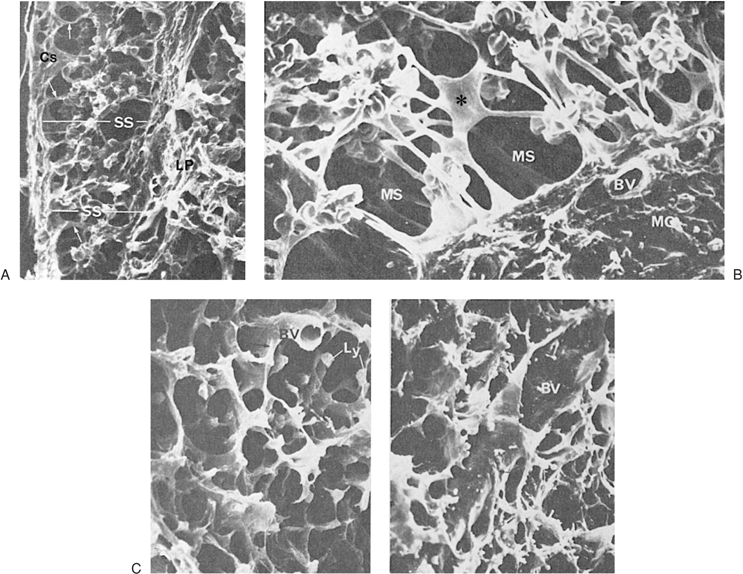
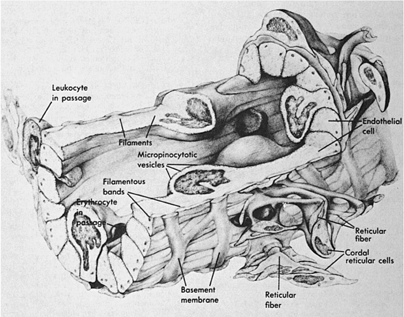
The basic framework of the LN is composed of trabeculae made of bundles of collagen and a few elastic fibers (123,124). This fibrous reticulum is argyrophilic when silver impregnation techniques are applied, forming a closely knit spongelike framework, with the fibers branching frequently and anastomosing freely with each other. The fibers penetrate dense lymphatic tissue and also surround the sinuses. They are ensheathed by two kinds of cells (Fig. 14.11). On the side that borders the dense lymphoid tissue, the fibrous reticulum is covered by thin, elongated cells that have been identified as fibroblasts or fibroblastic reticulum cells. They have large nuclei and endoplasmic reticulum in their cytoplasm. Cytoplasmic processes join similar processes of adjacent cells and form a continuous cellular sheath (cellular reticulum), which separates the fibrous reticulum from the dense lymphoid tissue. On the side bordering the sinuses, the fibers are covered by flat cells containing multiple vesicles. These are the littoral or endothelial cells lining the sinuses, and have no basement membrane. Fibers sometimes cross the sinuses and are covered by endothelial cells (Fig. 14.12). They divide the lumen into smaller interconnecting compartments. These trabeculae not only provide mechanical support for the sinuses, but also slow the flow of the lymph, enhancing the opportunities for phagocytosis by macrophages present along their course or lining the sinus walls. The sinuses originate from a foramen on the inner wall of the subcapsular sinus where afferent lymphatics terminate. The outer walls of the subcapsular sinuses are formed by the capsule of the LN. The sinuses follow the fibrous trabeculae through the LN parenchyma. Viewed from this perspective, the reticulum divides the LN into two distinct spaces. One consists of conventional vascular channels, the sinuses, which are sparsely occupied by cells and have a rapid rate of flow. This compartment of the LN facilitates lymphocyte traffic. The other division may be regarded as vascular spaces with unique characteristics: They are lined by fibroblasts, are tightly packed with lymphocytes and other cells, and have slow rates of flow. Immune responses take place in this compartment through cellular interactions. Both of these spaces communicate through pores in the inner wall of the subcapsular sinus.
|
|
|
Figure 14.11. Ultrastructure of a lymph node. The area in the center and to the right is occupied by dense lymphoid tissue with many lymphoid cells. It is separated from the sinus (upper area, left, and below) by flat endothelial cells lining the sinus (s), bulky reticular cells (R) bordering the dense lymphoid tissue, and collagenous fibers (c) between the two cellular layers. Arrows indicate collagenous fibers traversing the dense lymphoid tissue. Cap, blood capillary. (From Clark SL Jr. The reticulum of lymph nodes in mice studied with the electron microscope. Am J Anat 1962;110:217, with permission.) |
Cellular Compartments
The most abundant type of cell in the LN is the lymphocyte. In the region beneath the capsule, known as the cortex, the lymphocytes are arranged in clusters known as follicles. If the follicle is composed of uniform small lymphocytes, it is a primary follicle; if it contains pale, lightly stained, blastlike cells with a euchromatic nucleus at its center, it is a secondary follicle. The central zone of the secondary follicle is called a germinal center and is the hallmark of antigenic stimulation. In the secondary follicles, the lymphocytes that surround the GC constitute the mantle or crescent. These germinal centers were so named by Flemming in 1885 because he considered them “breeding grounds for the generation of lymphocytes” (125). Later, Hellman called them “reaction centers,” important in the induction of immunity (126). In the areas between the follicles, lymphocytes are distributed diffusely (diffuse cortex). The region that separates the cortex from the medulla is the deep cortex or paracortex and contains tightly packed predominantly T lymphocytes.
The histologic arrangement of lymphocytes in the LN corresponds to distinct functions performed by separate classes of lymphocytes (Fig. 14.13). The follicles are occupied by B lymphocytes and represent the sites of intense activity during humoral (antibody) responses (see Chapters 15 and 17). Two types of lymphoid cells are present in the GC: The centroblasts (noncleaved), which are large and activated B lymphocytes, and the centrocytes (cleaved), which are small and are derived from the centroblasts. The centroblasts are located at the bottom of the GC (dark zone), and the centrocytes are located at the upper part (apical zone). The role of the GC in the maturation of the antibody response is discussed in Chapter 15. A subpopulation of centroblasts expresses a phenotype characteristic of Burkitt cells (127). GCs also contain a subset of T cells, which are CD4+, CD45RO+, and CD57+ (128), and belong to the Th2 subset. These T cells are important in initiating T–B cell interactions for antibody formation. A large number of B lymphocytes not selected for antibody synthesis (or generation of memory cells) die by apoptosis and are disposed of by phagocytic macrophages known as tingible body macrophages.
The mantle of the follicle is composed of small lymphocytes that morphologically appear identical. However, they are functionally and phenotypically heterogeneous. A small number are small lymphocytes not yet antigenically stimulated that have arrived from the bone marrow, are immunologically competent, and express both IgM and IgD. The majority are small, recirculating, and indistinguishable from the previous set but may have responded to T-independent antigens. A third population consists of memory B cells, which are long lived and recirculating, and express IgG or IgA. Some of the B lymphocytes of the mantle are CD5+. The relationship of this lymphocyte to CD5- B cells is discussed in Chapter 15.
The paracortex and diffuse cortex constitute the T-cell compartment that undergoes histologic changes during cell-mediated immunity. Sometimes in the paracortex, nodules consisting of interdigitating DCs and T cells are formed after an immune response (129).
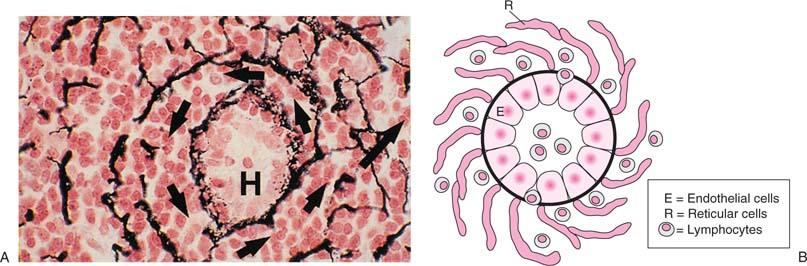
Cross sections of paracortex stained with special stains show a set of concentric rings, which form barriers or walls separating vascular spaces in between, where the lymph flows (Fig. 14.14). This structure constitutes the paracortical cord (130) with its center in the high endothelial venule (HEV) (see description later in this section). Lymphocytes exit the HEV through an interendothelial route and emerge into the perivenular channels (PVCs) (134), which is the space between the abluminal side of the endothelium and the surrounding pericytes. These channels are bounded by pericytes or fibroblastic reticular cells (FRCs). FRCs circumscribe the HEV with at least two layers of overlapping cells. Flow of lymphocytes through PVCs is rate limiting because the PVC around the HEV is extremely narrow, and lymphocytes must distend the channel to flow through it. On emergence from PVCs, the lymphocytes enter a space packed densely with cells, known as the corridor. The corridors are wide and can accommodate two lymphocytes side by side, and their walls are reticular fibers, enclosed by FRCs. The FRCs leave a narrow space surrounding the fibers, which is called the conduit. The corridors allow cellular interactions between slowly moving T cells that sample antigens on stationary DCs, “shaking hands much like the receiving line at a wedding” (131). The conduit, on the other hand, is a specialized system where fluid carries soluble factors important for the immune functions of the lymphocytes. The corridor space, where flow is slow, is separated from the sinuses, where flow is fast, by the reticular fibers and the endothelium of the sinuses.
In the medulla, the cells are arranged in cords (medullary cords) composed of lymphocytes, macrophages, and plasma cells.
|
|
|
Figure 14.12. Ultrastructure of a lymph node. A: Subcapsular sinus (SS) traversed by trabeculae (arrows) stretches between the capsule (Cs) and the parenchyma (LP) (×600). B: Medullary sinus (MS) with medullary cord (MC) and a meshwork of trabeculae (asterisk). (×1,900). C: Lymphoid parenchyma and the meshwork of trabeculae (arrows) separating intercommunicating cavernous spaces normally occupied by lymphocytes (Ly). (left, ×1,100; right, ×1,600). BV, blood vessel. (From Luk SC, Nopajarooysri C, and Simon GT. The architecture of the normal lymph node and hemolymph node. A scanning and transmission electron microscopic study. Lab Invest 1973;29:258, with permission.) |
|
|
|
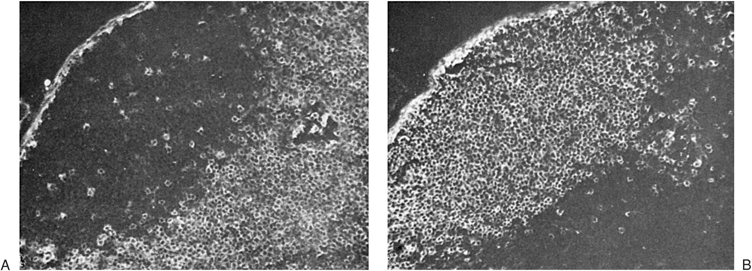
Figure 14.13. B- and T-cell–dependent areas of the lymph node. Frozen section of a mouse lymph node was examined by immunofluorescence with an anti-immunoglobulin antiserum (A) or an antiserum to T cells (B). A: Only the follicle fluoresces (B-cell–dependent area). B: The paracortex fluoresces brightly (T-cell–dependent area), whereas the follicle remains dark except for a few scattered fluorescent T cells. These are CD4+ helper T cells, which migrate to the germinal center because they express the chemokine receptor CXCR5 and are attracted to its ligand, BCA-1 chemokine released from cells in the follicle (see Lymphocyte Homing and Recirculation). (From Weissman IL, Weiryke R, Butcher EC, et al. The lymphoid system. Its normal architecture and the potential for understanding the system through the study of lymphoproliferative diseases. Hum Pathol 1978;9:25–45 with permission.) |
|
|
|
Figure 14.14. Cross section of a paracortical cord showing the corridor, outlined by reticular fibers stained by Gomori stain. A: The corridor encircles the high endothelial venule in the center (H). B: The drawing represents an “idealized” cross section of panel A. (Courtesy of Drs. Gretz, Anderson, and Shaw. From Gretz JE, Anderson AO, Shaw S. Cords, channels, corridors, and conduits: critical architectural elements facilitating cell interactions in the lymph node cortex. Immunol Rev 1997;156:11–24, with permission.) |
Other Cells: Dendritic Cells
The functions of lymphocytes are predicated on their interactions with other cells, such as macrophages and DCs.
Macrophages are found in large numbers in the walls of the sinuses and in the dense lymphoid regions. They phagocytose and rapidly remove foreign substances. Within minutes after injection, foreign substances are found in the phagolysosomes of medullary macrophages. Macrophages therefore contribute in an important way to the filtering functions of LNs as they capture and destroy infectious agents. In the process of phagocytosing foreign material, macrophages process antigen for presentation to lymphocytes, especially the T lymphocytes.
Antigen presentation, however, is mediated by other nonphagocytic professional antigen-presenting cells, generally known as dendritic cells(132). Their name vividly describes their morphology, with long arborizing cytoplasmic processes or dendrites (from the Greek dendron, “tree”), extending from the cell body for long distances. Within the LN, there are two distinct populations of DCs: One resides in the follicles, known as a follicular DC or FDC, and the second in the paracortex, known as interdigitating DC or IDC. Their origins, functions, and cellular associations are distinct. FDCs are autochthonous, whereas IDCs are immigrants that arrive from remote parts of the body using the routes of the lymphatic vessels. The FDCs of the follicles have an irregularly shaped nucleus with long, beaded cytoplasmic processes. They are not phagocytic but retain antigen for prolonged periods, captured in the form of immune complexes, and function as arbiters of life and death for emerging centrocytes based on high or low affinity, respectively, of their receptor for antigen (see Chapters 15 and 17). They also have been implicated in the generation of memory lymphocytes and the maintenance of antibody production. Two types of FDCs have been described, one with filiform and one with beaded dendrites (133). The FDCs interact with B lymphocytes through adhesion molecules such as intercellular adhesion molecule-1 (ICAM-1) (CD54) and vascular cell adhesion molecule-1 (VCAM-1) (CD106), which interact with the ligands LFA-1 (CD11a/CD18 or αLβ2integrin) and VLA-4 (CD49d), respectively, on the surface of the B cells (134). The FDC network may be disrupted in angioimmunoblastic lymphadenopathy, acquired immunodeficiency syndrome, and Hodgkin disease (135).
Electron microscopic studies of the primary follicles detected reticulum cells, with little or no evidence of phagocytic activity, making contacts with neighboring cells by very tight adhesions or desmosomes (136). These “reticulum cells” are stellate in shape, have long cytoplasmic processes, and are probably of mesenchymal origin. GC FDCs probably derive from the primary follicle reticulum cells. Dendritic reticulum cells do not ensheathe collagenous fibers and therefore are distinct from the fibroblastic reticulum cells that form and ensheathe the collagenous fibers.
The IDCs are located in the paracortex and extend numerous cytoplasmic processes that surround T cells. The IDC is not phagocytic but carries antigen from the periphery to present it to T cells in the paracortex of the LNs. Tight junctions between IDCs and T lymphocytes have been detected by electron microscopy and become particularly prominent after stimulation with antigens that elicit strong cell-mediated immunity. Their cytoplasm is electron lucent and contains lysosomelike vesicles and tubular structures extending to the periphery of the cell. The IDCs surrounded by clusters of T cells form the paracortical nodules around endothelial venules, important in the generation of antigen-specific T lymphocytes. T-cell nodules are detected next to B-cell follicles, forming the “composite nodules.” Prominent T-cell nodules are detected in dermatopathic lymphadenitis, where large numbers of LCs migrate from the areas of inflamed skin. Similar changes are also seen in regional LNs draining areas of T-cell lymphomas involving the skin. Mechanisms and routes of migration of IDCs are discussed in the section Dendritic Cell Migration.
In ontogeny the dendritic system is slow in developing and as a result during neonatal life the body depends primarily on innate immunity (137).
Vasculature
Ontogeny
Growth factors of the vascular endothelial growth factor (VEGF) family control lymphangiogenesis. They activate VEGF-specific receptors of which VEGFR-3 (Flt4), VEGF-C, and VEGF-D have been implicated in the regulation of lymphatic development and lymphangiogenesis. Both are processed after synthesis and form mature dimers of the VEGF domain. Both bind with high affinity to VEGFR-2 and VEGFR-3 expressed on endothelial cells. VEGFR-3 is a receptor tyrosine kinase with seven extracellular Ig-like domains and cytoplasmic tyrosine kinase domain (138,139). Growth of the endothelial cells in the high endothelial venules is regulated by CD11c+ dendritic cells secreting VEGF (140). This function optimizes the microenvironment for an immune response.
|
|
|
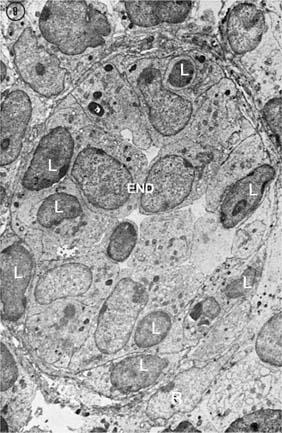
Figure 14.15. A postcapillary venule is practically occluded by the tall endothelial cells (End). Some lymphocytes (L) are crossing the wall of the high endothelial venule and enter the lymph node. The wall is surrounded by reticular cells (R) (×4,500). (From Clark SJL Jr. The reticulum of lymph nodes in mice studied with the electron microscope. Am J Anat 1962;110:217, with permission.) |
Lymphatic Vessels
Afferent lymphatics pierce the capsule and empty into subcapsular sinuses. Cortical sinuses originate from the subcapsular sinus and often run along the trabeculae and through the cortex to the medulla, where they become medullary sinuses. Eventually, the sinuses give rise to the efferent lymphatic vessels that leave the LN at the hilum.
Blood Vessels
The artery enters the LN at the hilum, gives rise to arterioles that reach the cortex along the trabeculae, and finally split into a rich network of capillaries. The capillaries empty into venules that extend from the cortex to the medullary cords and eventually exit from the hilum as veins. The venules linked most immediately to the capillaries possess unique endothelial cells that are tall and cuboidal and therefore contrast with the endothelial linings of other venules that usually are low or flat. The HEVs (Figs. 14.15 and 14.16) are the sites at which lymphocytes migrate from the blood circulation into the lymphatic circulation. Lymphotoxin signaling controls the expression of addressing an HEV, especially GleNAc-6-0 sulfotransferase-1 and -2. As a result, homing to the paracortex is drastically reduced, resulting in a marked reduction of the cellularity of the lymph node (141). The high endothelial cells have abundant cytoplasm, and their luminal surface is covered by a 1.5-nm-thick coat composed of filamentous and granular material. On the basis of the content of their polyribosomes, two types of cells are distinguished by electron microscopy; one is lighter and one darker, and the latter is also intensely pyroninophilic. The cytoplasm also contains numerous microtubules that radiate from the centriole, as well as multiple dense bodies that probably are related to lysosomes but also may function as storage sites for glycoproteins used in the formation of the cell coat. One striking morphologic feature is the large Golgi complex with myriad vesicles, a characteristic associated more commonly with actively secreting cells than with endothelial cells. Indeed, these cells secrete chemokines, which direct transendothelial migration of lymphocytes in HEVs (see section Lymphocyte Homing and Recirculation).
The bulky nucleus protrudes into the lumen and contains a well-developed reticulated nucleolus, characteristics of a metabolically highly active cell. Acid hydrolase activity and nonspecific esterases that are not present in other endothelia have been detected in these cells, as well as a peculiar ability to shift to anaerobic metabolism.
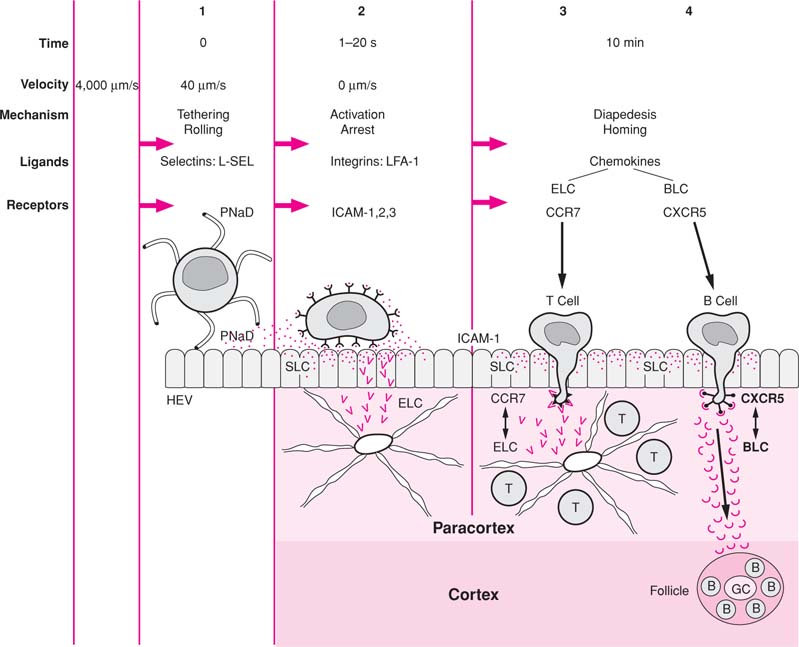
The height of the endothelial cells and their metabolic activity are influenced by the number of circulating T lymphocytes. Neonatal thymectomy and chronic thoracic duct drainage in mice and rats result in flattening of the endothelium, loss of pyroninophilia, and absence of lymphocytes migrating through the HEV wall (142). Infusion of lymphocytes into these animals reconstitutes the normal morphologic appearance of HEVs. In congenitally athymic (nude) mice and rats, the endothelium of HEVs is flat. Some evidence also has implicated the macrophages as playing a regulatory role in determining the morphology and metabolic activity of the endothelial cells (143). The HEVs are surrounded by layers of sheaths that derive from cytoplasmic plates of reticular cells linked to the fibrous reticulum of the LN. These sheaths may regulate the passage of lymphocytes to the LN while limiting fluid leakage and providing vascular support (Fig. 14.14B). When Marchesi and Gowans (144) originally identified the HEVs as the site for lymphocyte migration, they thought that lymphocytes cross the HEV wall by penetrating the cytoplasm of the high endothelial cells. This crossing is now recognized as an artifact because all lymphocytes that appeared on electron microscopic sections to be surrounded by endothelial cells were in fact outside of these cells when evaluated with tracer studies. Thus, the lymphocytes migrate across the HEV wall by insinuating themselves between endothelial cells (145,146). The interactions of lymphocytes and the HEV during homing and recirculation are described in the section Transendothelial Migration.
Morphologic Changes During the Immune Response
Antigens enter the LN via the afferent lymphatics that empty into the subcapsular sinus. Antigen then percolates in the sinuses through the parenchyma and is eventually found in two areas of the LN. In the medulla, it is localized within the lysosomes of macrophages (147), whereas in the cortex, it is associated with the membranes of FDCs (148). Antibody facilitates this follicular localization. During a humoral response, intense proliferative activity is observed in the center of the follicles 1 to 2 days after immunization. By 72 hours, large pyroninophilic cells are seen in the GCs of draining LNs (149). These cells have been given various names, such as immunoblasts, a term introduced originally by Dameshek (150), or germinoblasts. To avoid confusion with gonadal cells, however, they were renamed centroblasts. This blast is a large cell (larger than 10 μm) and has a nucleus with loose chromatin and a giant reticulated nucleolus that may occupy as much as one half of the nuclear area. The Golgi apparatus is moderately developed and contains many free polyribosomes but only few cisternae of endoplasmic reticulum. As the endoplasmic reticulum becomes more abundant, these cells are known as plasmablasts (151). At this stage, their nucleus possesses clumps of heterochromatin and a moderate-sized nucleolus. Free polyribosomes are still present, and the cisternae of the endoplasmic reticulum do not fill the cytoplasm entirely. By day 5, an increase is noted in the number of tingible body macrophages that give the GCs the appearance of a starry sky. Tingible body macrophages (Korper of Flemming) are macrophages that have phagocytosed nuclear debris of lymphocytes (the graveyard theory) (152). Because of their intense basophilic cytoplasm, the immunoblasts (or centroblasts) demarcate a histologic area known as the dark zone (also known as the basalor lower zone because it is located in the area of the GC that is toward the interior of the LN) (153). As they become more mature (immunocytes, centrocytes), they occupy the light zone (also called the upper zone because it is located above the dark zone toward the capsule). Small numbers of plasma cells are detected in the GCs.
|
|
|
Figure 14.16. Lymphocytes crossing the high endothelial venule (HEV). Lymphocyte migration is shown in successive stages (1,2,3,4). After attachment to the luminal surface of an endothelial cell (1), the lymphocytes insinuate between endothelial cells (2) and cross the basement membrane (3) and a sheath of reticular cells (RC) surrounding the HEV (4). The endothelial cells are attached to the neighboring cells by junctional complexes and to the perivascular sheaths by foot processes (FP). (From Anderson AO, Anderson ND. Lymphocyte emigration from high endothelial venules in rat lymph nodes. Immunology 1976;31:455, with permission.) |
Kinetic studies with 3H-thymidine provided evidence of the origin of centrocytes from the centroblasts and the dissemination of the activated cells beyond the confines of the GC (154). Little doubt remains that the GC blasts originate from B lymphocytes, and display a unique phenotype characterized by lack of membrane IgD. The formation of GCs depends on the presence of T lymphocytes (155).
The significance of the GC reaction is discussed in Chapter 17. They are the source of plasma cells (156), seen in the medullary cords by day 3 or 4 after immunization, as well as memory cells. For antigens that stimulate strong cell-mediated immunity, the proliferative activity and increase in the pyroninophilic blast cells are noted in the paracortex, which is the T-cell–dependent area.
Functions
LNs should be viewed as dynamic structures, the cells of which are constantly on the move. The functions of LNs can be considered in two broad categories. As an efficient mechanical filter, the LN includes an exquisite arrangement of the reticular meshwork and the sinuses with strategically located master phagocytic cells, the macrophages. If the filtration function fails to control infectious agents, the LNs become new foci for spread of disease.
The LNs also constitute important sites of immune functions that depend on complex interactions between lymphocytes and other cells. To this effect, LNs provide a suitable microenvironment, which includes the HEVs that permit lymphocytes to enter (home) and recirculate; stromal cells that direct lymphocytes to distinct domains, such as the IDCs of the paracortex that attract T lymphocytes and present them antigens; and the FDCs in the follicles that interact with B cells. LNs thus are important sites for the generation of effector mechanisms, including antibodies and T-cell–mediated immune responses. Both responses are exported from LNs and are carried to remote sites to control infections.
Spleen
The orphan homeobox gene 11 (Hox11) controls the expression of the spleen (157). Hox-1 knockout mice lack the spleen but otherwise appear normal. No defects in other secondary lymphoid organs or bone marrow and thymus have been detected.
|
|
|
Figure 14.17. Drawing of the splenic structure and circulation. PALS, periarteriolar lymphatic sheath. |
Although the spleen is considered a secondary lymphoid organ, it serves several other functions unrelated to the immune system. The spleen affects all blood cells throughout their lifespan (158). It provides the microenvironment for the final differentiation of reticulocytes, platelets, and monocytes; it removes aged or deformed red blood cells; and it is a reservoir for erythrocytes and granulocytes.
Structure
The spleen is covered by a connective tissue capsule that is several millimeters thick (159). The human spleen contains only a few muscle cells and thus is not capable of marked contractions. The internal surface of the capsule is the point of origin for an extensive network of trabeculae several millimeters in each dimension that divides the organ into communicating compartments. The appearance is spongelike, with the spaces containing the parenchyma or pulp tissue.
The capsule is indented on the medial surface where the blood vessels, lymphatics, and nerves enter and leave. Sections of the spleen stained with hematoxylin and eosin reveal two kinds of parenchyma: One is stained dark blue to purple because of the predominance of small lymphocytes (the white pulp) and the second is red because of the predominance of blood-filled sinuses (the red pulp) (Fig. 14.17).
White Pulp
The white pulp is the main lymphatic tissue of the spleen and surrounds the blood vessels (Figs. 14.19 and 14.20). The splenic artery branches into trabecular arteries that follow the trabeculae and in turn give off branches that leave the trabeculae. These structures are now called central arteries because of their central position in the white pulp (Fig. 14.20). They are muscular vessels that can alter blood flow by their contraction and relaxation. The white pulp is composed of the supporting reticulum, the free cells, and the blood vessels.
The reticulum is a scaffolding made of reticular fibers produced by reticular cells. The reticulum cells are spindle shaped with conspicuous microfilaments in their cytoplasm and prominent endoplasmic reticulum. The nucleus contains finely dispersed chromatin. This reticulum provides support for the free cells and the blood vessels. It runs circumferentially around the central artery and becomes particularly pronounced in the periphery of the white pulp, where the reticular cells extend sheets of membranes concentrically (160,161) (Fig. 14.19A).
|
|
|
Figure 14.18. Section of the spleen. White pulp (WP): Lymphocytes (L) packed around an arteriole (A). Red pulp (RP) surrounds the white pulp and consists mainly of sinuses, the cords, and cordal spaces. |
The central artery gives off secondary branches that radiate through the white pulp with different destinations. Some vessels terminate before they reach the edges of the white pulp, and some go beyond and terminate in the marginal zone (MZ) or extend even further into the red pulp. In the human spleen, some branches curve back from the red pulp and form a delicate network around lymphatic follicles located at the periphery of the white pulp.
|
|
|
Figure 14.19. Ultrastructure of the spleen by scanning electron microscope. A: Periarteriolar lymphatic sheath with a central artery (A). Tightly packed lymphocytes lie between fibers and reticular cells (arrows) that run circumferentially around the artery. B: Lumen of a sinus at upper left has elongated endothelial lining cells. The cord below contains bulky reticular cells (asterisks) and fibers (white arrows). Some erythrocytes (E) are visible in the cordal spaces. The large cell in the lumen of the sinus is probably a macrophage with several thin cytoplasmic strands (arrow). The beaded black lines identify at the cut edge of the sinus its adventitial layer (terminal bead) and its endothelial layer (subterminal bead). C: Lumen of a sinus that bifurcates to the right. The elongated endothelial cells are in close apposition without any visible gaps. (From Weiss L. A scanning electron microscopic study of the spleen. Blood 1974;43:665, with permission.) |
The predominant cell type of the white pulp is the lymphocyte, which occupies the interstices of the spongelike meshwork of the reticulum, forming two histologically distinct arrangements. Immediately surrounding the central artery, lymphocytes are tightly packed in a diffuse sheath that follows the central artery much like a cellular cylinder, eventually becoming thinner as the artery divides into small arterioles. This diffuse lymphocytic collection is known as the periarteriolar lymphatic sheath or PALS (Figs. 14.18 and 14.19A). The PALS is divided into the central and peripheral areas. The central area immediately surrounding the artery is occupied by tightly packed T lymphocytes. The stroma in this area contains the interdigitating cells seen also in the T-dependent area (paracortex) of the LN (162). The PALS in the human spleen is not continuous but at intervals the arteriole runs across a follicle without T cells, but re-enters again a T-cell region (163). T lymphocytes usually are found in intimate contact with the interdigitating cells. The periphery of PALS lacks the interdigitating cells and contains B and T lymphocytes. Lymphocyte collections in the form of follicles are seen in this region, some of them containing blastlike cells at their center that form the GC (secondary follicle). The small lymphocytes that surround the GCs constitute the mantle that separates the GC from the MZ. These GCs, like their counterparts in the LN, can be divided histologically into a dark zone occupied by large pyroninophilic blasts with high mitotic activity and a light zone with more mature cells. The light zone, which faces the MZ, has an abundance of the characteristic FDCs that also are present in the nodules of the LN. They are large cells with unusually shaped nuclei that are poor in heterochromatin, and the cytoplasmic extensions are thin sheets rather than dendrites. The FDCs are not phagocytic but retain foreign substances on their membranes for long periods. No FDC can be located in the central section of PALS and their presence in the mantle of the nodules facing the MZ places them in a strategic location to capture foreign substances entering the spleen. Antigens are localized first in the MZ and then cross the marginal sinus to lodge in the mantle of the nodules, forming a crescentic cap. The nodules of the peripheral section of PALS constitute the B-dependent regions of the spleen. The segregation of the two major lymphocyte populations into distinct settlements within the secondary lymphoid organs seems to be determined by the nature of their underlying neighbors.
|
|
|
Figure 14.20. A human splenic sinus. The elongated lining cells run parallel to the axis of the sinus. The vessel is supported by cordal reticular cells and fibers and a fenestrated basement membrane consisting of heavy fibrous strands running circumferentially and lighter longitudinal strands. Thus, the circular bands, like the hoops of a barrel, hold the endothelial cells (staves of the barrel) tightly. (From Weiss L. The spleen. In: Weiss L, ed. Cell and tissue biology. Baltimore: Urban and Schwarzenberg, 1988, with permission.) |
Marginal Zone
The MZ is the part of the parenchyma that lies between the white and red pulps (Figs. 14.17 and 14.18). This area of the spleen is important because it comes in contact with large quantities of circulating blood. Many arterial vessels terminate in this region, some of them in funnel-shaped orifices that empty their contents into the interstices of the marginal zone. Unlike the red pulp, the MZ has no sinuses. In rodents, a marginal sinus separates the MZ from the white pulp, but the human spleen lacks a marginal sinus. Some authors suggest that the marginal sinus may be functionally equivalent to the postcapillary venule and, in rodents, the site at which lymphocytes enter the splenic parenchyma. The lymphocytes entering the marginal sinus have multiple microvilli with which they establish contacts with reticular cells as they find their way toward their microenvironments. Once they reach their destination, the microvilli disappear. However, the marginal sinus does not have the tall endothelial cells that direct traffic in the LNs. Some authors speculate that this function is performed by splenic macrophages located in the marginal sinus. These cells have distinct phagocytic and morphologic properties that distinguish them from macrophages in other locations of the spleen, including the ability to bind lymphocytes (164). Splenic processing of blood cells begins in the MZ.
Marginal Zone Lymphocytes: Functions and Malignancies
In human spleen, the MZ contains clusters of B cells and macrophages in the vicinity of the periphery of the white pulp. Morphologically, phenotypically, and genotypically, these cells differ from naïve B cells. They are large with pale cytoplasm and irregular nucleus. They are IgD-, CD5-, CD10-, and CD23-, and have mutated their Ig-V genes (165,166). MZ B cells are memory cells, but constitute a heterogeneous population in response to antigens and VH gene rearrangements.
In terms of response to antigens, there are three types, those responding to T-cell–dependent antigens, and those responding to T-cell–independent antigens, which are of two types: Type 1 (i.e., lipopolysaccharides [167]) or type 2 (i.e., bacterial capsular polysaccharides [165]).
B cells are born in the bone marrow and when they move out to the lymphoid organs they are subject to maturation that gives rise to two morphologically and functionally distinct subsets: (a) the follicular B cells and (b) the MZ B cells (168). The follicular B cells are IgMlow and IgDhigh, are small in size and recirculating, and reside in the lymph node follicles. The MZ B cells are medium-size IgMhigh and IgDlow and reside in MZ of the spleen. On the basis of cell-surface markers, it was suggested that maturation of B cells in the periphery proceeds through three transitional stages and the MZ B cell arises from an intermediate stage with the CD21/CD35high, CD23+ phenotype (169). Furthermore, they are able to respond rapidly to certain antigens filtered from the blood (168). In addition to the morphologic and phenotypic difference between follicular and MZ B cells, there are also differences in their mechanisms of activation and signaling pathways, such as the use of extracellular signaling molecule Notch 2 (170), as well as in their migration (171) and retention (172).
An intriguing and important concept for the development of MZ B cells has been suggested from data showing that the MZ B-cell development depends on signal strength (173) from self-antigens and is independent of exogenous antigens (174). Some B cells in lymph nodes have a similar phenotype and morphology as the MZ B cells and are known as nodal-MZ B cells or monocytoid B cells. The MZ B cells and nodal MZ B cells rearrange the same VH3 family of genes (suggesting that both of these B-cell populations are closely related) (175). Others, however, brought evidence that the monocytoid B cells are distinct from the MZ B cells, on the basis of their phenotype (i.e., lack of IgM and IgD expression from most of them, polyclonal Ig gene rearrangements with lack of somatic mutations) (176). However, whether hypermutation takes place in the MZ B cells has been challenged, because the activation-induced cytidine deaminase (AID) (see Chapter 17), the enzyme that is essential for somatic hypermutation and class-switch recombination, is not detected in MZ B cells (177). This find raises the question as to how an MZ B cell subpopulation mutates their Ig genes. One possibility is that MZ B cells may recognize self-lipids presented by CD1 of NK T cells (178).
Another intriguing finding that may shed some light on the obscure nature of the MZ B cells and their origin is a recent finding that the BCR of the MZ B cells is of a polyreactive nature including self-activity, such as against single-stranded DNA (179). In spite of intensive research, the nature of the MZ B cell in humans is not yet clear. Results from experimental animals may not necessarily apply to humans (180). Expression of CD27+ B cells in human spleens is detected in the periphery of the follicles corresponding to the mantle, but it is not spleen specific. It most likely is associated with memory B cells (181).
B-cell neoplasms involving a cell with strikingly similar morphologic, phenotypic, and cytogenetic characteristics have been reported to have different anatomic localizations (splenic, nodal, and extranodal) and clinical presentations (182,183).
Variations detected by mutational analysis were considered as evidence that these lymphomas originate from different subsets of marginal cells (184). More recent studies of MZ B-cell lymphomas utilizing complementary DNA microarray analysis identified expression of three genes that, according to this study, may distinguish splenic MZ lymphoma: ILF1, senatasin, and CD40 (185). Furthermore, the evidence that mucosal-associated lymphomas preferentially disseminate to the spleen was considered as evidence of the relationship of these cells to the splenic MZ cell (186).
Red Pulp
On a histologic section, the red pulp consists of sinuses, which give the pulp its color, and cords (187,188). The cords are composed of a meshwork of reticular fibers and cells. The reticular cells are large and extend membranous processes into the interstices (Figs. 14.17, 14.18, and 14.19) that contact those of adjacent cells, forming cavernous spaces. The large reticular cell sheets have microfilaments that endow them with the capacity to retract and extend, thus determining the available space and regulating blood flow. These cordal spaces receive blood directly from the arterial vessels. The central artery gives off many branches and terminates in slender, straight, nonanastomosing arterioles that enter the cords of the red pulp, not the sinuses. Some of the arterioles divide into arterial capillaries that are enveloped by a sheath of phagocytic cells.
These sheaths were called ellipsoids, but now are called periarterial macrophage sheaths. In the human spleen, the periarterial macrophage sheath is not well developed. As well as functioning as a major source of phagocytic cells, the periarterial macrophage sheath also may regulate blood flow. The cords have been considered to represent a unique vascular space, the flow through which is regulated by the cordal reticular cells. In addition, removal of old or damaged red cells take place in the cords.
The red pulp sinuses are 35 to 40 μm in diameter and are tortuous vascular channels lined with an endothelium consisting of elongated, tapered cells arranged with their long axes parallel to that of the vessel (Fig. 14.19B,C). The endothelial cells have three distinctive morphologic features: (a) micropinocytotic vesicles, (b) loosely organized cytoplasmic filaments, and (c) tightly organized filaments along the basal side, which provide the cell with rigidity and contractility. Slitlike gaps between the endothelial cells allow the penetration of blood cells from the cordal spaces into the lumen. The slits never exceed 0.2 to 0.5 μm in width. The intraendothelial filaments run parallel to the slits and restrict their opening (189). They arch between adjacent ring components of the basement membrane. Although normal red blood cells are flexible and capable of passing through the slits, the presence of rigid inclusions, such as Heinz bodies, interferes with their passage. The basement membrane is not continuous but consists of a network of reticular fibers that resemble a roll of chicken wire. Fibers running circumferentially (annular fibers) are thick and are joined by thinner fibers that run longitudinally (Fig. 14.20). The network is connected to the reticulum of the cords. Some reticular cells are in close contact with the wall of the sinuses. Through the fenestrations of this network, blood cells pass from the cords to the sinuses. Adventitial cells cover the sinus wall from the cordal side. They contribute to blood flow regulation by covering or exposing the interendothelial slits.
The splenic circulation has been the subject of debate. According to some investigators, the circulation is closed, with endothelial continuity from the arteries to the sinuses and veins. Others believe the circulation is open—that the arteries empty their blood into the MZ and red pulp cords and the blood percolates through the cavernous spaces of the cords and finally crosses the wall of the sinuses through the interendothelial slits. According to this view, the circulation is anatomically open but is functionally closed. This question has not been resolved, but it is likely that both types of pathways exist. In keeping with this last hypothesis is the finding that the largest amount of blood passes through the spleen as rapidly as it does through other organs, whereas a smaller portion of blood has a considerably slower rate of flow.
Morphologic Changes during the Immune Response
Within 24 hours of antigenic stimulation, an accumulation of polymorphonuclear leukocytes is detected in the red pulp (151). Large pyroninophilic blast cells are first seen 3 to 4 days later in the PALS. They then migrate toward the periphery as they mature to plasma cells. They eventually reach the cords of the red pulp and are found even within the sinuses. GCs are well developed by the end of the first week. When viewed by electron microscopy, the most immature blasts have a large nucleus, a prominent nucleolus, and an increase in the number of ribosomes, but no organized endoplasmic reticulum is evident. These blasts are known as immunoblasts, or plasmablasts, if they develop dilated endoplasmic reticulum. Proplasmacytes are the next stage of maturation. These cells have abundant endoplasmic reticulum and a few remaining free ribosomes, findings that differentiate this stage of development from mature plasma cells.
Immunologic Significance of the Spleen
The two most important immunologic functions of the spleen are phagocytosis and the development of the effector mechanisms of humoral and cell-mediated immunity (190). The labyrinthine structure of the spleen is ideal for the effective removal of invading microorganisms by phagocytes that come to reside in the spaces of the reticulum. Phagocytosis is facilitated by the coating of bacteria with complement, antibody, or both. Although the phagocytic function of the spleen is well recognized, its contribution to humoral or cell-mediated immunity in humans has generated considerable controversy (191,192). Originally, it was reported that children who underwent splenectomy for congenital hemolytic anemia had an increased incidence of overwhelming, fatal infections, but not all subsequent studies yielded results that confirmed these findings. Splenectomy for hematologic disorders apparently has far more serious consequences than splenectomy for trauma. The incidence of overwhelming sepsis in the former cases is 24.8% as compared to a 1.45% incidence for the latter group (193). The contribution of the spleen to humoral responses in laboratory animals has been known for a long time. In humans, the loss of the spleen also compromises the immune system and its ability to form antibody (194). Even impaired antibody synthesis by peripheral blood lymphocytes has been detected in postsplenectomy patients (195). In spleenless Hox-/- mice, there is no major defect in antiviral immunity except for some delay in antibody response by 1 to 2 days (196).
![]() Lymphocyte Homing and Recirculation
Lymphocyte Homing and Recirculation
Homing to Secondary Lymphoid Organs
Migration of leukocytes across the vascular endothelium is a complex, multistep process and involves different types of endothelia for different leukocytes, as well as a variety of adhesion molecules.
When B and T lymphocytes finish their maturation in the bone marrow and thymus, respectively (primary lymphoid organs), they are still naïve—that is, they have not yet encountered antigenic stimulation. The migration patterns of naïve lymphocytes differ from those of the activated (or memory) lymphocytes. Naïve lymphocytes enter the secondary lymphoid organs and settle in specific (T-dependent and T-independent) compartments (homing). Subsequently, they use the blood and lymphatic circulation as traffic routes and migrate between secondary lymphoid organs without any specific preference (recirculation). Antigen-activated or memory lymphocytes migrate specifically to sites where they have encountered antigen. Thus, memory T lymphocytes tends to accumulate in extranodal tissues where they have previously been antigenically stimulated. These lymphoid accumulations are sometimes called tertiary lymphoid tissues and are associated with epithelial surfaces such as the gut, respiratory tree, and sites of inflammation in skin and synovium. In some of these accumulations, however, there are also small numbers of naïve lymphocytes.
If antigen is drained to regional LNs, it triggers accumulation of antigen-specific memory T lymphocytes to these nodes. In general, two fundamentally distinct routes of entry of lymphocytes to the LN can be distinguished. The first is by crossing of the HEV, which is primarily followed by the naïve lymphocytes, and the second is by the afferent lymphatics after the lymphocytes have entered tissue spaces by crossing flat endothelium. The second route is followed by the activated T lymphocytes (197).
Homing of lymphocytes to secondary lymphoid organs takes place through the HEVs, which are abundant in T-cell–dependent areas of the LN, but are used as entry sites for both T and B lymphocytes (198,199,200,201). In the spleen, homing takes place through blood sinusoids in the MZ. HEVs are also found in other extranodal locations, such as PPs, tonsils, adenoids of the pharynx, appendix, and other lymphocyte aggregates of the stomach and small intestine.
|
|
|
Figure 14.21. Homing of T and B lymphocytes to peripheral lymph nodes. 1. Rolling: Lymphocytes entering the high endothelial venule (HEV) slow down as a result of interaction of the L-selectin (L-SEL) (lymphocytes) with its ligand the peripheral node addressin (PNad) on endothelial cells. 2. Arrest: Secondary lymphoid tissue chemokine (SLC) chemokine, constitutively produced by HEV cells, activates integrin αLβ2 (CD11a/CD18) and tightly adheres lymphocyte to the endothelial surface. 3. Diapedesis: SLC (produced by HEV cells) and Epstein-Barr virus–induced molecule 1 ligand chemokine (ELC) (produced by dendritic cells and trancytosed to the surface) act on appropriate receptors (CCR7) and attract T (stronger) and B (weaker) cells that insinuate themselves through the endothelial cells. 4. Home settling: Entering the node, T cells settle in the paracortex (ELC production), while B cells and a few antigenically experienced CD4+ T cells move on, being attracted by another chemokine, B lymphocyte chemoattractant (BLC) (or BCA-1), acting on both of them, expressing the receptor CXCR5 to settle in the follicles. Gc, germinal center; ICAM, intercellular adhesion molecule; INT, integrin. |
The best HEV marker currently available is a carbohydrate epitope recognized by the monoclonal antibody MECA-79 (202).
Crossing of HEVs by lymphocytes involves three steps, which have been defined for interactions of all leukocytes with endothelia: (a) tethering and rolling, (b) integrin activation and arrest or tight adhesion, and (c) transendothelial migration (Fig. 14.21).
Tethering and Rolling
Tethering is the establishment of loose and transient contacts between the lymphocyte and the endothelial cell. This adhesion is weak, and the shear forces of blood flow result in the rolling of the lymphocyte along the endothelium. Tethering to and rolling along the vascular endothelium, in this case the HEV, is mediated by L-selectin (CD62L, previously known as LECAM-1, MEL14, and LAM-1) on lymphocytes and its counterreceptors on HEV cells.
For L-selectin to mediate rolling, it must be located on the tips of the microvilli (203,204). The L-selectin mediates the primary adhesion (rolling) on HEVs of peripheral LNs. However, for homing of naïve lymphocytes to the secondary lymphoid organs of the gut (PPs), L-selectin requires the cooperation of the integrin α4β7, which is not required for peripheral LN homing. This integrin is important for activated T lymphocytes that lack L-selectin expression (205). The importance of L-selectin for lymphocyte adhesion to HEVs was shown with blocking antibodies and by gene knockout, which decreases the lymphocyte migration to LN by 99% (206). Rolling by L-selectin requires linkage to cytoskeletal proteins such as α-actinin, vinculin, and talin through its 11 carboxy terminal amino acids (207). Rolling enables leukocytes to reduce speed and thus be able to sense chemoattractants in order to activate firm adhesion. The tether bond dissociation increases exponentially as shear stress increases, but this is compensated for with an increase in the number of selectin–ligand bonds between the lymphocyte and the endothelial cell, while at the same time, the bonds are clustered and collectively resist breaking more efficiently than individually. At higher shear, the average lifetime of rolling tethers is much longer as a result of elongation of microvillous tethers. This type of kinetics stabilizes leukocyte rolling over a wide range of shear forces (208).
It is accepted that a ligand for L-selectin needs to carry three critical structural elements: Sialic acid, fucose, and sulfate. Monoclonal antibodies specific for sialyl 6-sulfo Lewis X oligosaccharide stain HEVs and almost completely inhibit binding of L-selectin to human HEVs. Cotransfection of complementary DNAs for 6-sulfotransferase and α1,3 fucosyl transferase VII (Fuc-TVII) generates significant L-selectin binding. L-selectin, however, prefers that this ligand is sulfate at carbohydrate 6 (209).
Antigens that carry L-selectin–binding saccharides are known as vascular addressins. One is known as glycosylation cell adhesion molecule 1 or GlyCAM1, and is a secreted protein not found on the cell membrane, which argues against a direct function as a cell adhesion molecule. In mice deficient in the GlyCAM1 gene, lymphocyte trafficking still occurs.
A second antigen is CD34, detected on endothelial cells throughout the vasculature, therefore not HEV specific. Furthermore, disruption of the CD34 gene does not abolish trafficking through the HEV.
At the present time, the nature of the macromolecule that carries the epitope detected by the monoclonal antibody MECA79, which defines an HEV-specific carbohydrate epitope, remains elusive and is known as peripheral node addressin or PNad (210,211). Biochemical and genetic evidence infers that, for the synthesis of sulfated glycan moieties, sulfation precedes the 2S3 sialylation, which in turn would be followed by α1,3fucosylation, required for an L-selectin ligand. With targeted deletion of FucT-VII locus, which encodes an α1,3 fucosyltransferase expressed in HEVs, there is an 80 to 90% reduction of short-term lymphocyte homing. And yet, some other work suggested that although sulfate on the epitope is required for L-selectin binding, neither sialic acid (212) nor fucose (213) is directly involved in the binding.
Sulfate groups are required for optimal L-selection binding, because if the sialyl Lewis X (i.e., sialyl-α[2-3]–galactopyranosyl-β[1-4]–N-acetylglucosamine-[α(1,2,3)-fuco-pyranosyl]) is modified with sulfate, it generates a critically important L-selectin recognition determinant. The most important sulfate modification is known as “sialyl 6-sulfo Lewis X” (sialyl Lewis X with sulfation of GlcNAc on the sixth position) (212). Sulfation is catalyzed by GlcNAc-6-0-sulfotransferases (GlcNAc6STs) with five members being identified (214,215). L-selectin–mediated leukocyte rolling is extremely fast and jerky in nature (216). This is probably due to endothelial chemokines juxtaposed to endothelial L-selectin ligands (217). The rolling leukocyte is probably detached from its binding site and binds to another ligand downstream, causing the jerky nature of rolling. The endothelial chemokines CCR7 and CXCK5 are G-protein–coupled receptors (GPCRs) or SLC/ELC and BCA-1, respectively. It is hypothesized that clusters of GPCR around L-selectin prevent the association of multiple L-selectin molecules necessary for stabilization of L-selectin contacts.
The sulfated/sialylated-O-linked oligosaccharides recognized by L-selectin are displayed on sialomucin core proteins. One of them is endomucin, which is expressed in HEVs of the lymph nodes but also in endothelial cells of other vessels. One of the sialomucins is endomucin, decorated with carbohydrate chains reactive with L-selectin. The endomucin isoform a is expressed on PNad of HEVs. Appropriately modified by glycotransferases and sulfotransferases, it can function as a ligand for L-selectin (218). Endomucin isolated from human and mouse lymphoid organs carried the MECA-79 epitope (219).
The P-selectin glycoprotein ligand-1 (DSGL-1), a mucinlike glycoprotein, is detected on various leukocytes and is a ligand for P-selectin, but it also binds to L-selectin and E-selectin. The enzymes fucosyltransferase (FucT)-IV and FucT-VII confer the L-selectin binding activity (220). This is another molecular mechanism for the transendothelial migration of lymphocytes as well as other leukocytes.
Integrin Activation and Arrest
In preparation for transendothelial migration, lymphocytes need to adhere firmly on the endothelial cells. Arrest is achieved by activation of LFA-1 (αLβ2) integrin, which interacts with ICAM-1 or ICAM-2 (204). On resting lymphocytes, LFA-1 exists in an inactive form but within <1 second, it is activated by chemokines constitutively expressed on lymphoid tissues (“lymphoid chemokines”) (221,222,223), such as secondary lymphoid tissue chemokine (SLC, or CCL21) (224,225) and Epstein-Barr virus–induced molecule 1 ligand chemokine (ELC, or CCL19), also called macrophage inflammatory protein-3β (MIP-3β) (226). SLC induces arrest of naïve T cells to ICAM-1 (227) with sixfold higher efficiency for naïve than memory T cells (228). The arrest is pertussis toxin sensitive, implying that the signaling mechanism is mediated through G proteins. The ELC shares the same binding specificity with SLC, but its role is less obvious. ELC transcripts are restricted to nonendothelial cells in the perivascular area of paracortex (229). However, it is brought to the luminal surface of the HEV by transcytosis and is able to participate in LFA-1 activation and arrest of the lymphocytes through the same receptor CCR7 as SLC (230). The integrin activation and affinity is regulated by conformational changes of their extracellular domains (231). These changes occur through extension of the genu, followed by separation of the stalks that lead to the high-affinity conformation (see details of integrin structure and activation in Chapter 17).
Transendothelial Migration
SLC induces strong chemotaxis for T cells and to a lesser extent on B cells (224). Entry of lymphocytes to the secondary lymphoid organs, especially T cells, requires the expression of CCR7, the high-affinity receptor for its ligand SLC. SLC is very important for entry of T cells into secondary lymphoid organs, as shown in mice with the plt mutation (paucity of LN T cells). These mice lack SLC lymphocytes, cells cannot enter through HEVs, and the structure of the paracortical areas is severely disorganized. The SLC gene in mice is located on chromosome 4 (232), which corresponds to human chromosome 9p13, where SLC is located. Furthermore, T cells lacking the SLC receptor (CCR7) have a markedly reduced ability to enter LNs and PPs.
In CCR7-deficient mice, B-cell homing into secondary lymphoid organs is not disturbed. The arrest of B cells on HEVs takes place in the absence of SLC, indicating that other chemotactic activities regulate entry of B cells into secondary LNs. SDF-1 is recognized by the CXCR4 receptor, expressed on resting T and B cells. This chemokine/receptor system may also contribute to lymphocyte entry into LNs. However, this interaction is not important for T cells as CXCR4-deficient mice have normal T-cell areas. The redundancy of the mechanisms for B-cell homing explains the normal B-cell homing in plt mice, as well as on SDF-1 and CXCR4-deficient mice. Transgenic mice that do not produce SLC on the normal anatomic locations but have SLC expression directed to pancreas under the control of the insulin promoter II have collections of CD4+ and CD8+ cells near the center of the islets and only a few scattered B cells surrounding them (233). In older mice, these clusters in the islets develop stromal reticulum, seen in LNs and vascular endothelium with morphology of HEVs. B-cell follicularlike clusters without GCs merge with the T-cell areas. This study clearly demonstrates the important role of SLC in LN neogenesis.
Once naïve lymphocytes have crossed the HEV, chemokines direct the lymphocytes for compartmental homing (i.e., T lymphocytes to the paracortex and B lymphocytes to the follicles). SLC is expressed not only on HEVs, but also by stromal cells within the T-cell areas of LNs, spleen, and PPs. A second ligand for CCR7 is ELC, which is also expressed in T-cell areas and is made by macrophages and the DCs of the paracortex. In vitro SLC and ELC attract T cells effectively, but B cells only weakly.
In conclusion (Fig. 14.21), T cells cross the HEV by the interaction of SLC and ELC chemokines with their receptor CCR7. B cells cross the HEV by more than one chemokine/receptor system. However, once inside the LN, they are attracted to the follicles by chemokine CXCL13 (BLC, BCA-1, B-cell–attracting chemokine 1), which binds to the receptor CXCR5 (or BLR1 [Burkitt lymphoma receptor 1]). The CXCR5 is expressed by mature as well as activated B cells and in addition by CD4+-activated T cells. Antigen-activated CD4+ T cells down-regulate CCR7 (receptor for SLC and ELC) and up-regulate CXCR5. Therefore, this CD4+ T-cell subset, in contrast to naïve T cells that stay in the paracortex, travels with B cells to the follicles. This explains the early fluorescent antibody results, which detected small numbers of scattered T cells within the GC (Fig. 14.13A). They induce strong antibody responses in culture with B cells (234).
CXCR5-deficient mice show defective follicular localization of B cells. BLC is expressed constitutively by FDC in lymphoid follicles of LNs and other secondary lymphoid organs, and in BLC-deficient mice there are severe defects in the development of secondary lymphoid organs. Histologically, B cells do not appear organized in follicles but form a ring around the T-cell areas (235). The importance of BLC for organization of B cells into follicles was shown by the ectopic expression of BLC in pancreatic islets where B lymphocytes but not T cells accumulate in these sites (236). Organization of primary follicles requires the network of FDCs, which in turn depends on LTα1β2 and TNF expressed on B cells. BLC, for its action, is strongly dependent on these cytokines (237).
In conclusion, for proper formation of follicles and localization of B lymphocytes, two ligand-receptor systems are mandatory: The TNF/LTα1β2ligand and its receptor LT βR, and the BLC ligand and its receptor CXCR5.
Signaling in Lymphocyte Homing
The Ras-homologous A protein (Rho A), a guanosine triphosphate–binding protein, is involved in the intracellular signaling for the integrin activation pathway (238,239). RhoA activation depends on the cytoplasmic tail of ICAM-1 but the exact mechanism is not known. The mechanism of coupling to the Ras/Map kinase cascade is not yet well defined. Activation of Ras leads to stimulation of a cascade of protein kinases that ultimately phosphorylate and activate mitogen-activated protein kinase. Phospholipase Cβ activation results in the production of inositol triphosphate and diacylglycerol. Inositol triphosphate is important in mobilizing Ca2+ from intracellular sources, whereas diacylglycerol is important in the activation of protein kinase C. PI3-kinase is also activated in response to chemokine stimulation, which links further downstream with phospholipase D activation, actin rearrangements, and vesicular trafficking (240,241). The role of PI3-kinase in lymphocyte homing is not clear, although PI-3 kinase is activated by chemokines (241). There are several PI3-kinase isoforms and cytokines may alter the balance between them and thus affect cell migration. PI3-kinase also cooperates with DOCK2 and optimal migration of T cells depends on expression of both (242).
DOCK2, a mammalian homologue of the Caenorhabditis elegans CED-5 protein family, activates Rac, which in turn regulates actin cytoskeleton and therefore cell mobility. DOCK2 mediates migration of lymphocytes independently of PI3-kinase.
The intracellular signals result in reorganization of the cytoskeleton, causing cells to send out pseudopodia and move along the chemoattractant gradient.
Transendothelial migration occurs rapidly, and the lymphocyte reaches the basement membrane within minutes. The cell moves toward the cell–cell junctions. Adhesive contacts with the endothelium are modulated so that the cell is not immobilized. Modulation may involve decrease of activation signals.
Dendritic Cell Migration
The traffic of DCs is regulated by cytokines (IL-1, TNF-α), chemokines, and β-defensins. Immature DCs, regardless of their origin, express the chemokine receptor 6 (CCR6), which binds the chemokine MIP-3α (CCL20) (243). In the case of LCs (244), it directs them to the skin, where the ligand for CCR6 (i.e., MIP-3α) is constitutively expressed by keratinocytes, which provide the homing signals for the positioning of LCs to the suprabasal layer of epidermis. β-Defensins produced by mucosal membranes after microbial infection bind to CCR6 and thus attract immature DCs to submucosal areas (245).
LCs are connected physically with keratinocytes through homotypic interactions mediated by E-cadherin. Inflammatory or allergic stimuli down-regulate E-cadherin, resulting in “maturation” and detachment of LCs (246). CD40-CD40 ligand may also play a role for their emigration (247). Detached LCs down-regulate CCR6 and up-regulate another chemokine receptor, CCR7, which allows the now mobile LCs to move toward the lymphatic vessels in response to its ligand, SLC (248). The barrier to their migration, imposed by the basement membrane of the skin (collagen IV), is brought down through the matrix metalloproteinase-9, released by LCs with specificity to collagen IV (249). LCs cross the abluminal side of the lymphatic vessels, probably with the involvement of P-glycoprotein, and eventually enter LNs. In the lymphatic vessels, they are known morphologically as veiled cells on the basis of their morphology (i.e., large lamellipodia [veils], which, like sails, propel them forward). They enter the LN from the afferent lymphatic vessel into the subcapsular sinus to reach the T-cell areas, attracted by SLC and MIP-3β (or ELC), both of them ligands of CCR7 receptor (250). In the T-cell areas, they are known as IDCs, as their elaborate long cytoplasmic processes interdigitate between T cells. IDCs present to T-cell antigens they have captured in the periphery, initiating an immune response (see Chapter 17).
Special Homing Pathways
Skin Homing
Memory lymphocytes tend to return to the type of tissue where they have encountered the antigen (251). Lymphocytes that entered the afferent lymphatics from the skin have a distinct phenotype (252). T lymphocytes localized in the skin express a marker called cutaneous lymphocyte-associated antigen (CLA), which binds to E-selectin (253,254). E-selectin is induced on dermal endothelial cells in the delayed type of hypersensitivity and in chronically inflamed skin (255). Other molecules also mediate strong adhesion of memory T cells to skin, such as α4β1(VLA-4) interacting with VCAM-1, and the integrin α3β1 with the extracellular matrix component epiligrin (256). However, T lymphocytes may penetrate the epidermis using E-cadherin.
Homing to the Gut
Lymphoid cells in the gut are found in three areas: As well-organized collections known as Peyer patches, which are secondary lymphoid organs; in the lamina propria underlying the intestinal epithelium; and after penetration of the basement membrane interspersed between epithelial cells—that is, the intraepithelial lymphocytes (IELs).
Naïve lymphocytes home to PPs, whereas memory cells home to extralymphoid sites—that is, the intestinal lamina propria as a result of variations in the expression of highly specialized homing receptors. The HEV of the PPs is weak for ligand of L-selectin and MAdCAM-1hi, whereas the naïve lymphocytes are L-selectin+ and α4β7lo–med. Therefore, neither set of molecules alone is capable of mediating tethering and rolling to initiate integrin activation, but cooperation between the two sets of molecules allows naïve lymphocytes to enter the PPs. The endothelium of the lamina propria is negative for L-selectin ligand, but strong for MAdCAM-1 expression. Naïve lymphocytes are unable to use L-selectin and, being weak in α4β7 expression, they cannot initiate rolling and are blocked from entering the lamina propria. On the other hand, memory lymphocytes, although they lack L-selectin (and as a result cannot enter PPs), have strong α4β7 expression and are able to initiate rolling and enter the lamina propria.
Some of the lymphocytes are able to penetrate the basement membrane of the intestinal epithelium and settle between enterocytes to become IELs. IELs interact with intestinal epithelium through an integrin known as the human mucosal lymphocyte molecule, now shown to be the αEβ7integrin (257,258). αEβ7 is expressed in approximately 2 to 6% of blood T cells, which are a memory subset and do not express CLA1 and L-selectin (259). The αEβ7 integrin is induced by transforming growth factor-β (260). The same cytokine also is known to trigger class switch of IgM+ B cells to IgA, which is the predominant Ig class produced by the gut lymphocytes. It is of interest that MAdCAM-1, a gut mucosal addressin, contains an IgA-like domain (261).
Homing to Inflammatory Sites
Lymphocytes are known to accumulate in sites of chronic inflammation and tissues with active autoimmune mechanisms. The phenotype of these lymphocytes indicates that they are activated, and therefore antigenically stimulated (251). Cytokines produced from macrophages, such as interferon-γ and TNF-α, regulate lymphocyte traffic to inflammatory sites. Accumulation of lymphocytes at chronic inflammation sites involves several adhesion molecules such as LFA-1 and α4 integrins, as well as E-selectin and VCAM-1. Chemoattractants, such as MIP-1β, enhance binding of CD8+ T cells (262), whereas RANTES attracts selectively memory T cells (263). The monocyte chemoattractant protein-1 (MPC-1) is a major lymphocyte chemoattractant (in addition to its strong activity on monocytes) (216); this explains the mixed composition of cellular infiltrates, rich in lymphocytes and monocytes, in sites of chronic and autoimmune inflammatory reactions.
MCP-1 is present in abundance in areas of antigenic challenge and autoimmune diseases (264). The data suggest that -C-C chemokines provide the triggering signal for the integrin-dependent tight adhesion of lymphocytes to endothelia for emigration to inflammatory sites.
Homing to Other Sites
Hemopoietic progenitors in bone marrow transplantation use the α4β1–VCAM-1 set to home to bone marrow (265). Interaction of P-selectin with its ligand, P-selectin–dependent ligand (PSGL-1), is also implicated (266). Lymphocyte homing to tonsils is mediated by L-selectin and CD34 (267), whereas the attachment of lymphocytes to brain microvasculature is mediated by the sulfoglucuronosyl paragloboside, a ligand for L-selectin (268).
Significance of Lymphocyte Homing and Recirculation
It is estimated that as many as 5 × 106 lymphocytes/second leave the blood and cross HEVs in the body. The process of rolling and sticking to the HEV takes only 1 to 20 seconds and the transendothelial passage approximately 10 minutes (269). From in situ microscopic observations of PPs, the velocity of the lymphocyte before contact with the HEV is approximately 4,000 μm/second, and rolling effectively reduces this to approximately 40 μm/second (206). Thus, rolling increases the chances for firm adhesion, but at the same time the cell may escape if proper triggering for activation of integrin mechanisms fails to be initiated. Triggering mechanisms involve G-protein–linked chemoattractant receptors and their ligands, and they can bring about integrin activation within 1 to 3 seconds after contact for tight adhesion.
Homing and recirculation are not random, but the result of active interactions determined by the phenotypes of the HEV cells and the lymphocytes. Homing to specific microenvironments of the secondary lymphoid organs is developmentally determined (i.e., T dependent vs. T independent), and carried out under the guidance of chemokines and their receptors and the cellular composition of the microenvironments (i.e., IDCs vs. FDCs) (270). When lymphocytes are antigenically stimulated, the pattern of migration, known as recirculation, is modified as a result of this experience. Antigenically specific lymphocytes are directed to particular tissues so that they may influence more effectively the intensity of the immune response. This ultimately is the significance of lymphocyte recirculation as reflected in the difference between naïve lymphocytes and the lymphocytes that become antigenically experienced. Homing and recirculation are forms of lymphocyte differentiation. These, like many other aspects of their differentiation (i.e., IgM vs. IgG production or Tho vs. Th1 or Th2 cytokine pattern secretion), are refined in the periphery in response to antigenic stimulation so that they best meet the environmental challenges for the protection of the body. Immune responses within the secondary microenvironments (271,272) as well as “tertiary” microenvironments (i.e., the areas of disease and inflammation) are all under the control of the chemokines (261). Knowledge of the mechanisms of chemokine regulation will allow the development of therapeutic strategies to control inflammatory diseases (273).
References
1. Marschalko T von. Ueber die sogenannten plasmazellen; ein beitrag zur kenntniss der herkunft der entzundlichen infiltrationszellen. Arch Derm Syph 1895;30:241.
2. Downey H. The origin and structure of the plasma cells of normal vertebrates, especially of the cold blooded vertebrates, and the eosinophils of the lung of Amblystoma. Folia Haematol 1911;11:275.
3. Maximow A. Untersuchungen uber blut und bindegewebe. VIII. Die zytologischen eigenschaften der fibroblasten, reticulumzellen und lymphozyten des lymphoiden gewebes ausserhalb des organismus ihre genetischen wechselbeziehungen und prospektiven entwicklungspotenzen. Arch Mikroskop Anat 1923;97:283.
4. Bloom W. Relationships between lymphocytes, monocytes and plasma cells. Folia Haematol (Leipz) 1928;37:63.
5. Rich AR. Inflammation in resistance to infection. Arch Pathol 1936;22: 228–254.
6. McMaster PD, Hudack SS. The formation of agglutinins within lymph nodes. J Exp Med 1935;61:783–805.
7. Dougherty TF, Chase JH, White A. The demonstration of antibodies in lymphocytes. Proc Soc Exp Biol Med 1944;57:295–298.
8. Harris TN, Grimm E, Mertens E, et al. The role of the lymphocyte in antibody formation. J Exp Med 1945;81:73–83.
9. Ehrich WE, Harris TN. The site of antibody formation. Science 1945;101: 28–31.
10. Fagraeus A. Antibody production in relation to the development of plasma cells. In vivo and in vitro experiments. Acta Med Scand 1948;204 (Suppl):1–122.
11. Coons AH, Leduc EH, Conolly JM. Studies on antibody production. I. A method for the histochemical demonstration of specific antibody and its application to a study of the hyperimmune rabbit. J Exp Med 1955;102:49–61.
12. Leduc EH, Coons AH, Conolly JM. Studies on antibody production. II. The primary and secondary responses in the popliteal lymph node of the rabbit. J Exp Med 1955;102:61–82.
13. Gudat FG, Harris TN, Harris S, et al. Studies on antibody producing cells. I. Ultrastructure of 19s and 7s antibody-producing cells. J Exp Med 1970;132: 448–474.
14. Harris TN, Hummeler K, Harris S. Electron microscopic observations on antibody producing lymph node cells. J Exp Med 1966;123:161–172.
15. Hummeller K, Harrris TN, Tomasini N, et al. Electron microscopic observations on antibody producing cells in lymph and blood. J Exp Med 1966;124:255–262.
16. Nowell PC. Phytohemagglutinin: an initiator of mitosis in cultures of normal leukocytes. Cancer Res 1960;20:462–466.
17. Bessis M. Living blood cells and their ultrastructure. New York: Springer-Verlag, 1973.
18. Grossi CE, Greaves MF. Normal lymphocytes. In: Zucker-Franklin D, ed. Atlas of blood cells. Function and pathology, 2nd ed. Philadelphia: Lea & Febiger, 1988.
19. Bessis M, Thiery JP. Electron microscopy of human white blood cells and their stem cells. Int Rev Cytol 1961;12:199–241.
20. Polliack A, Lampen N, Clarkson BD, et al. Identification of human B and T lymphocytes by scanning electron microscopy. J Exp Med 1973;138:607–624.
21. Dwyer JM. Identifying and enumerating human T and B lymphocytes: a review of techniques, problems, and progress in clinical studies. Prog Allergy 1976; 21:178–260.
22. Lewis WH. Locomotion of lymphocytes. Bull Johns Hopkins Hosp 1931;49: 29–36.
23. McFarland W, Heilman DH, Moorhead JE. Functional anatomy of the lymphocyte in immunological reactions in vitro. J Exp Med 1966;124:851–858.
24. McFarland W, Schechter GP. The lymphocyte in immunological reactions in vitro: ultrastructural studies. Blood 1970;35:683–688.
25. Werdelin O, Braendstrup O, Petersen E. Macrophage-lymphocyte clusters in the immune response to soluble protein antigen in vitro. I. Roles of lymphocytes and macrophages in cluster formation. J Exp Med 1974;140:1245–1259.
26. Schreiner GF, Unanue ER. Membrane and cytoplasmic changes in B lymphocytes induced by ligand-surface immunoglobulin interaction. Adv Immunol 1976;24:37–165.
27. Zucker-Franklin D. Structural features of cells associated with the paraproteinemias. Semin Hematol 1964;1:165.
28. Lichtman MA. The ultrastructure of the hemopoietic environment of the marrow: a review. Exp Hematol 1981;9:392–410.
29. Campioni D, Moretti S, Ferrari L, et al. Immunophenotypic heterogeneity of bone marrow-derived mesenchymal stromal cells from patients with hematologic disorders: correlation with bone marrow microenvironment. Hema- tologica 2006;91:364–368.
30. Taichman RS. Blood and bone: two tissues whose fates are intertwined to create the hematopoietic stem-cell niche. Blood 2005;105:2631–2639.
31. Taichman RS, Reilly MJ, Emerson SG. Human osteoblasts support human hematopoietic progenitor cells in in vitro bone marrow cultures. Blood 1996;87:518–524.
32. Whitlock CA, Witte ON. Long-term culture of B lymphocytes and their precursors from mucine bone marrow. Proc Nat Acad Sci U S A 1982;79:3608–3612.
33. Yoshida T, Ng SY-M, Zuinga-Pflucker JC, et al. Early hematopoietic lineage restrictions directed by Ikaros. Nat Immunol 2006;7:382–391.
34. Nagasawa T. Microenvironmental niches in the bone marrow required for B cell development. Nat Rev Immunol 2006;6:107–116.
35. Egawa T. The earliest stages of B cell development require a chemokine stromal cell derived factor/pre-B cell growth-stimulating factor. Immunity 2001;15:323–334.
36. Sitnicka E. Complementary signaling through flt3 and interleukin-7 receptor α is indispensable for fetal and adult B cell genesis. J Exp Med 2003;198:1495–1506.
37. Ma O, Jones D, Sprigner TA. The chemokine receptor CXCR4 is required for the retention of B lineage and granulocytic precursors within the bone marrow environment. Immunity 1999;10:463–471.
38. vanFreeden-Jeffrey V. Lymphopenia in interleukin (IL)-7 gene-deleted mice identifies IL-7 as a non-redundant cytokine. J Exp Med 1995;181:1519–1526.
39. Haynes BF. The human thymic microenvironment. Adv Immunol 1984;36:87.
40. Weiss L. The thymus. In: Weiss L, ed. Cell and tissue biology. Baltimore: Urban and Schwarzenberg, 1988.
41. Hollander G, Gill J, Zukleys S, et al. Cellular and molecular events during early thymus development. Immunol Rev 2006;209:28–46.
42. Cordier AC, Heremans JF. Nude mouse embryo = ectodermal nature of the primordial thymic defect. Second J Immunol 1975;4:193–196.
43. LeDouarin NM, Jotereua FV. Tracing of cells of the avian thymus through embryonic life in interspecific chimerals. J Exp Med 1975;142:17–40.
44. Blackburn CC, Manley NR. Developing a new paradigm fore thymus organogenesis. Nat Rev Immunol 2004;4:278–288.
45. Zou D, Silvius D, Davenport J, et al. Patterning of the third pharyngeal pouch into thymus/parathyroid by Six and Eya1. Dev Biol 2006;293:499–512.
46. Gordon J, Wilson VA, Blair NF, et al. Functional evidence of a single endodermal origin for the thymic epithelium. Nat Immunol 2004;5:546–553.
47. Nehls M, Pfeifer D, Schorpp M, et al. New member of the winged-helix protein family disrupted in mouse and rat nude mutations. Nature 1994;372:103–107.
48. Schuddekopf K, Schorpp M, Boehm T. The whn transcription factor encoded by the nude locus contains an evolutionarily conserved and functionally indispensable activation domain. Proc Natl Acad Sci U S A 1996;93:9661–9664.
49. Le Lièvre CS, Le Douarin NM. Mesenchymal derivatives of the neural crest: analysis of chimaeric quail and chicken embryos. J Embryol Exp Morphol 1975;34:125–154.
50. Patel DD, Hale LP, Whichard LP, et al. Expression of CD44 molecules and CD44 ligands during human thymic fetal development: expression of CD44 isoforms is developmentally regulated. Int Immunol 1995;7:277–286.
51. Hollander GA, Wong B, Nichogiannopoulou A, et al. Developmental control point in induction of thymic cortex regulated by a subpopulation of prothymocytes. Nature 1995;373:350–353.
52. Ritter MA, Boyd RL. Development in the thymus: it takes two to tango. Immunol Today 1993;14:462–469.
53. Muller KM, Leudecker CJ, Udey MC, et al. Involvement of E-cadherin in thymus organogenesis and thymocyte maturation. Immunity 1997;6:257–264.
54. Kutlesa S, Wessels JT, Speiser A, et al. E-cadherin-mediated interactions of thymic epithelial cells with CD103+ thymocytes lead to enhanced thymocyte cell proliferation. J Cell Sci 2002;115:4505–4515.
55. Anderson M, Anderson SK, Farr AG. Thymic vasculature: organizer of the medullary epithelial compartment? Int Immunol 2000;12:1105–1110.
56. Erickson M, Morkowski S, Lehar S, et al. Regulation of thymic epithelium by keratinocyte growth factor. Blood 2002;100:3269–3278.
57. Dunn RJ, Leudecker CJ, Haugen HS, et al. Thymic overexpression of CD40 ligand disrupts normal thymic epithelial organization. J Histochem Cytochem 1997;45:129–147.
58. Farr AG, Dooley JL, Erickson M. Organization of thymic medullary epithelial heterogeneity: implications for mechanisms of epithelial differentiation. Immunol Rev 2002;189:20–27.
59. Bjorses P, Aaltonen J, Horelli-Kuitunen N, et al. APECED: a new clue to autoimmunity. Hum Mol Genet 1998;7:1547–1553.
60. Kumar PG, Laloraya M, Wang C-Y, et al. The autoimmune regulator (AIRE) is a DNA-binding protein. J Biol Chem 2001;276:41357–41364.
61. Su MA, Anderson MS. Aire: an update. Curr Opin Immunol 2004;16:746–752.
62. Tao Y, Kupfer R, Stewart BJ, et al. AIRE recruits multiple transcriptional components to specific genomic regions through tethering to nuclear matrix. Mol Immunol 2006;43:335–345.
63. Pitkaenen J, Peterson P. Autoimmune regulator: from loss of function to autoimmunity. Genes Immun 2003;4:12–21.
64. Kogawa K, Nagafuchi S, Katsuta H, et al. Expression of AIRE gene in peripheral monocyte/dendritic cell lineage. Immunol Lett 2002;80:195–198.
65. Klein L, Kyewski B. “Promiscuous” expression of tissue antigens in the thymus: a key to T-cell tolerance and autoimmunity? J Mol Med 2002;78:483–494.
66. Anderson MS, Venanzi ES, Chen Z, et al. The cellular mechanism of Aire control of T cell tolerance. Immunity 2005;23:227–239.
67. Hughes AL, Friedman R. A cross-tissue expression and evolution of genes controlled by the Aire transcription factor. Genomics 2006;88:462–467.
68. Terszowski G, Muller SM, Bleul CC, et al. Evidence for a functional second thymus in mice. Science 2006;312:284–287.
69. Miller JF. Immunological function of the thymus. Lancet 1961;2:748–749.
70. Miller JFAP. Investigating a second thymus in mice. Science 2006;312: 1597–1598.
71. von Boehmer H. Thoracic thymus, exclusive no longer. Science 2006;312:206–207.
72. Wallin J, Eibel H, Neubuser A, et al. Pax1 is expressed during development of the thymus epithelium and is required for normal T-cell maturation. Development 1996;122:23–30.
73. Hetzer-Egger C, Scharpp M, Haas-Assenbaum A, et al. Thymopoiesis requires Pax9 function in thymic epithelial cells. Eur J Immunol 2002;32:1175–1181.
74. Dooley J, Erickson M, Roelink H, et al. Nude thymic rudiment lacking functional foxn1 resembles respiratory epithelium. Dev Dyn 2005;233:1605–1612.
75. Pantelouris DM. Absence of thymus in a mouse mutant. Nature 1968;217: 370–371.
76. Mecklenburg L, Nakamura M, Sundberg JP, et al. The nude mouse skin phenotype: the role of Foxn1 in hair follicle development and cycling. Exp Mol Pathol 2001;71:171–178.
77. Bleul CC, Boehm T. Chemokines define distinct microenvironments in the developing thymus. Eur J Immunol 2000;30:3171–3379.
78. Balcirunaite G, Kellar MP, Balcirunaite E, et al. Wnt glycoproteins regulate the expression of Foxn1, the gene defective in nude mice. Nat Immunol 2002;3:1102–1108.
79. Rossi SW, Jenkinson WE, Anderson G, et al. Clonal analysis reveals a common progenitor for thymic cortical and medullary epithelium. Nature 2006;441: 988–990.
80. Bleul CC, Corbeaux T, Reuter P, et al. Formation of a functional thymus initiated by a postnatal epithelial progenitor cell. Nature 2006;441:992–996.
81. van Ewijk W. Immunohistology of lymphoid and non-lymphoid cells in the thymus in relation to T lymphocyte differentiation. Am J Anat 1984;170:311–330.
82. Wekerle H, Ketelsen UP, Ernst M. Thymic nurse cells. Lymphoepithelial cell complexes in murine thymuses: morphological and serological characterization. J Exp Med 1980;151:925–944.
83. Hassal AH. The microscopic anatomy of the human body in health and disease. London: Samuel Highly Publications, 1849.
84. Lobach DF, Scearce RM, Haynes BF. The human thymic microenvironment: phenotypic characterization of Hassall’s bodies with the use of monoclonal antibodies. J Immunol 1985;134:250–257.
85. Watanabe N, Wang Y-H, Kyre Lee H, et al. Hassall’s corpuscles instruct dendritic cells to induce CD4+ CD25+ regulatory T cells in human thymus. Nature 2005;436:1181–1185.
86. Wu L, Li CL, Shortman K. Thymic dendritic cell precursors: relationship to the T lymphocyte lineage and phenotype of the dendritic cell progeny. J Exp Med 1996;184:903–911.
87. Shortman K, Vremec D, Corcoran LM, et al. The linkage between T-cell and dendritic cell development in the mouse thymus. Immunol Rev 1998;165: 39–46.
88. Spits H, Blom B, Jaleco AC, et al. Early stages in the development of human T, natural killer and thymic dendritic cells. Immunol Rev 1998;165:75–86.
89. Ardavin C, Martinez del Hoyo G, Martin P, et al. Origin and differentiation of dendritic cells. Trends Immunol 2001;22:691–700.
90. Savchenko AS, Hasegawa G, Naito M. Development and maturation of thymic dendritic cells during human entogeny. Cell Tissue Res 2006;325:455–460.
91. Ito H, Esashi E, Akiyama T, et al. IL-18 produced by thymic epithelial cells induces development of dendritic cells with CD11b in the fetal thymus. Intern Immunol 2006;18:1253–1263.
92. Watanabe N, Hanabuchi S, Soumeles V, et al. Human thymic stromal lymphopoietin promotes dendritic cell-mediated CD4+ T cell homeostatic expansion. Nat Immunol 2004;5:426–443.
93. Bearman RM, Bensch KG, Levine GD. The normal human thymic vasculature: an ultrastructural study. Anat Rec 1975;183:485–497.
94. Raviola E, Karnovsky MJ. Evidence of a blood-thymus barrier using electron-opaque tracers. J Exp Med 1972;136:466–498.
95. Nienwenhuis P, Stet RJ, Wagenaar JP, et al. The transcapsular route: a new way for (self-) antigens to bypass the blood-thymus barrier? Immunol Today 1988;9:372–375.
96. Steinmann GG. Changes in the human thymus during aging. Curr Top Pathol 1986;75:43–48.
97. Min H, Montecino-Rodriguez E, Dorshkind K. Reduction in the developmental age. J Immunol 2004;173:245–251.
98. Montecino-Rodriguez D, Min H, Dorshkind K. Reevaluating current models of thymic involution. Sem Immunol 2005;17:356–361.
99. Shanker A. Is thymus redundant after adulthood? Immunol Lett 2004;91:79–86.
100. Beverly LJ, Ascano JM, Capobianco AJ. Expression of JAGGED 1 in T-lymphocytes results in thymic involution by inducing apoptosis of thymic stromal epithelial cells. Genes Immun 2006;7:476–486.
101. Tridente G. Immunopathology of the human thymus. Semin Hematol 1985;22:56–67.
102. Fuchs S, Schmidt-Hopfeld I, Tridente G, et al. Thymic lymphocytes bear a surface antigen which cross-reacts with acetylcholine receptor. Nature 1980;287:162–164.
103. Kao I, Drachman DB. Thymic muscle cell bear acetylcholine receptors: possible relation to myasthenia gravis. Science 1977;195:74–75.
104. Okumura M, Miyoshi S, Fujii Y, et al. Clinical and functional significance of WHO classification of human thymic epithelial neoplasms. Am J Surg Pathol 2001;25:103–110.
105. Fu YX, Chaplin DD. Development and maturation of secondary lymphoid tissues. Ann Rev Immunol 1999;17:399–433.
106. Cupedo T, Kraal G, Megius RE. The role of CD45+ CD4+ CD3- cells in the lymphoid organ development. Immunol Rev 2002;189:41–50.
107. Drayton DL, Liao S, Mounzer RH, et al. Lymphoid organ development: from ontogeny to neogenesis. Nat Immunol 2006;7:344–353.
108. Sabin FR. The lymphatic system in human embryos, with a consideration of the morphology of the system as a whole. Am J Anat 1909;9:43–91.
109. Wigle JT, Harvey N, Detman M, et al. An essential role for Prox1 in the induction of the lymphatic endothelial cell phenotype. EMBO J 2002;21:1505–1513.
110. De Togne P, Goelner J, Ruddle NH, et al. Abnormal development of peripheral lymphoid organs in mice deficient in lymphotoxin. Science 1994;264: 703–707.
111. Ware CF, Van Arsdale TL, Crow PD, et al. The ligands and receptors of the lymphotoxin system. Curr Top Microbiol Immunol 1995;198:175–218.
112. Weih F, Cosemano J. Regulation of secondary lymphoid organ development by the nuclear factor-κB signal transduction pathway. Immunol Rev 2003;195:91–105.
113. Muller G, Lipp M. Concerted action of chemokine and lymphotoxin system in secondary lymphoid-organ development. Curr Opin Immunol 2003;15: 217–224.
114. Ohl L, Henning G, Krautwald S, et al. Cooperating mechanisms of CXCR5 and CCR7 in development and organization of secondary lymphoid organs. J Exp Med 2003;197:1199–1204.
115. Mebius RE, Rennert P, Weissman IL. Developing lymph nodes collect CD4+ CD3- LTβ+ cells that can differentiate to APC, NK cells, and follicular cells but not to T or B cells. Immunity 1997;7:493–504.
116. Mebius RE, Miyamoto I, Christensen J, et al. The fetal liver counterpart of adult common lymphoid progenitors gives rise to all lymphoid lineages, CD45+ CD4+ CD3- cells, as well as macrophages. J Immunol 2001;166:6593–6601.
117. Kurebayashi S, Ueda E, Sakaue M, et al. Retinoid-related orphan receptor gamma (ROR gamma) is essential for lymphoid organogenesis and controls apoptosis during thymopoiesis. Proc Natl Acad Sci U S A 2000;97:10132–10137.
118. Yokota Y, Mansowi A, Mori S, et al. Development of peripheral lymphoid organs and natural killer cells depends on the helix-loop-helix inhibitor Id2. Nature 1999;397:702–706.
119. Kim D, Mebius RE, MacMicking JD, et al. Regulation of peripheral lymph node genesis by the tumor necrosis factor family member TRANCE. J Exp Med 2000;192:1467–1478.
120. Dougall WC, Glaccum A, Charrier K, et al. RANK is essential for osteoclast and lymph node development. Genes Dev 1999;13:2412–2424.
121. Miyawaki S, Nakamura Y, Suzuka H, et al. A new mutation, aly, that induces a generalized lack of lymph nodes accompanied by immunodeficiency in mice. Eur J Immunol 1994;24:429–434.
122. Ansel KM, Ngo VN, Hyman PL, et al. A chemokine-driven positive feedback loop organizes lymphoid follicles. Nature 2000;406:309–314.
123. Clark SL Jr. The reticulum of lymph nodes in mice studied with the electron microscope. Am J Anat 1962;110:217–257.
124. Luk SC, Nopajaroonsri C, Simon GT. The architecture of the normal lymph node and hemolymph node. A scanning and transmission electron microscopic study. Lab Invest 1973;29:258–265.
125. Flemming W. Studien uber regeneration der Gewebe. I. Die zellvermehrung in den lymphdrusen and verwandten organen und ihr einfluss auf deren bau. Arch Mikrosk Anat 1884;24:50.
126. Hellman T. Studien uber das lymphoide gewebe. Die bedeutung der sekundarfollikel. Beitrage Pathol Anat Allgem Pathol 1921;68:333.
127. Gregory CD, Tursz T, Edwards CF, et al. Identification of a subset of normal B cells with a Burkitt’s lymphoma (BL)-like phenotype. J Immunol 1987;139:313–318.
128. Bowen MB, Butch AW, Parvin CA, et al. Germinal center T cells are distinct helper-inducer T cells. Hum Immunol 1991;31:67–75.
129. Inaba K, Witmer MD, Steinman RM. Clustering of dendritic cells, hyper T lymphocytes, and histocompatible B cells during primary antibody responses in vitro. J Exp Med 1984;160:858–876.
130. Kelly RH. Functional anatomy of lymph nodes: the paracortical cords. Int Arch Allergy Appl Immunol 1975;48:836–849.
131. Gretz JE, Anderson AO, Shaw S. Cords, channels, corridors and conduits: critical architectural elements facilitating cell interactions in the lymph node cortex. Immunol Rev 1997;156:11–24.
132. Bancereau J, Briere F, Cause C, et al. Immunobiology of dendritic cells. Annu Rev Immunol 2000;18:767–811.
133. Szakal AK, Kosco MH, Tew JG. Microanatomy of lymphoid tissue during humoral immune responses: structure function relationships. Annu Rev Immunol 1989;7:91–109.
134. Freedman AS, Munro JM, Rice GE, et al. Adhesion of human B cells to germinal centers in vitro involves VLA-4 and ICAM-1. Science 1990;249:1030–1033.
135. Alavaikko MJ, Hansmann ML, Nebendahl C, et al. Follicular dendritic cells in Hodgkin’s disease. Am J Clin Pathol 1991;95:194–200.
136. Cyster JG, Ansel KM, Ekland EH, et al. Follicular stromal cells and lymphocyte homing to follicles. Immunol Rev 2000;176:181–193.
137. Dakic A, Shao Q-x, D’Amico AD, et al. Development of the dendritic cell system during mouse ontogeny. J Immunol 2004;172:1018–1027.
138. Achen MG, Stacker SA. The vascular endothelial growth factor family: proteins which guide the development of the vasculature. Int J Exp Pathol 1998;79:255–265.
139. Lymboussaki A, Partanen TA, Olofsson B, et al. Expression of the vascular endothelial growth factor C receptor VEGRF-3 in lymphatic endothelium in skin and vascular tumors. Am J Pathol 1998;153:395–403.
140. Webster B, Ekland EH, Agle LM, et al. Regulation of lymph node vascular growth by dendritic cells. J Exp Med 2006;203:1903–1913.
141. Browing JL, Allaire N, Ngam-ek A, et al. Lymphotoxin-B receptor signaling is required for the homeostatic control of HEV differentiation and function. Immunity 2005;23:539–550.
142. Goldschneider I, McGregor DD. Migration of lymphocytes and thymocytes in the rat. I. The route of migration from blood to spleen and lymph nodes. J Exp Med 1968;127:155–168.
143. Hendriks HR, Eestermans IL. Disappearance and reappearance of high endothelial venules and immigrating lymphocytes in lymph nodes deprived of afferent lymphatic vessels: a possible regulatory role of macrophages in lymphocyte migration. Eur J Immunol 1983;13:663–669.
144. Marchesi VT, Gowans JL. The migration of lymphocytes through the endothelium of venules in lymph nodes: an electron microscope study. Proc R Soc (Biol) 1964;159:283–290.
145. Anderson AO, Anderson ND. Lymphocyte emigration from high endothelial venules in rat lymph nodes. Immunology 1976;31:731–748.
146. Schoefl GI. The migration of lymphocytes across the vascular endothelium in lymphoid tissue. A reexamination. J Exp Med 1972;136:568–588.
147. Nossal GJV, Abbot A, Mitchell J. Antigens in immunity. XIV. Electron microscopic radioautographic studies of antigen capture in the lymph node medulla. J Exp Med 1968;127:263–276.
148. Nossal GJV, Abbot A, Mitchell J, et al. Antigens in immunity. XV. Ultrastructural features of antigen capture in primary and secondary lymphoid follicles. J Exp Med 1968;127:277–290.
149. Stein H, Gerdes J, Mason DY. The normal and malignant germinal center. Clin Haematol 1982;11:531–559.
150. Dameshek W. “Immunoblasts” and immunocytes. An attempt at a functional nomenclature. Blood 1963;21:243–245.
151. Movat HZ, Fernando NVP. The fine structure of the lymphoid tissue during antibody formation. Exp Mol Pathol 1965;4:155–188.
152. Swartzendruber DC, Congdon CC. Electron microscope observations on tingible body macrophages in mouse spleen. J Cell Biol 1963;19:641–646.
153. Mori M, Ishi Y, Onoé T. Studies on the germinal center. I. Ultrastructural study of germinal centers in human lymph nodes with correspondence to the zonal differentiation. J Reticuloendothel Soc 1969;6:140–157.
154. Hanna MG Jr. An autoradiographic study of the germinal center in spleen white pulp during early intervals of the immune response. Lab Invest 1964;13:95–104.
155. Jacobson EB, Caporale LH, Thorbecke GJ. Effect of thymus cell injections on germinal center formation in lymphoid tissues in nude (thymusless) mice. Cell Immunol 1974;13:416–430.
156. Nossal GJV, Makela O. Autoradiographic studies on the immune response. I. The kinetics of plasma cell proliferation. J Exp Med 1962;115:209–230.
157. Roberts CWM, Shutter JR, Korsmeyer SJ. Hox11 controls the genesis of the spleen. Nature 1994;368:747–749.
158. Lewis SM. The spleen—mysteries solved and unresolved. Clin Haematol 1983;12:363–373.
159. Weiss L. The spleen. In: Weiss L, ed. Cell and tissue biology. Baltimore: Urban and Schwarzenberg, 1988.
160. Weiss L. A scanning electron microscopic study of the spleen. Blood 1974;43:665–691.
161. Weiss L. The structure of the normal spleen. Semin Haematol 1965;2:205–228.
162. Veerman AJP, van Ewijk W. White pulp compartments in the spleen and rats and mice. A light and electron microscopic study of lymphoid and non-lymphoid cell types in T- and B-areas. Cell Tissue Res 1975;156:417–441.
163. Steiniger B, Rú´ttinger L, Barth PJ. The three-dimensional structure of human splenic white pulp compartment. J Histochem Cytochem 2006;51:655–663.
164. Lyons AB, Parish CR. Are murine marginal-zone macrophages the splenic white pulp analog of high endothelial venules? Eur J Immunol 1995;25:3165–3172.
165. Spencer J, Perry ME, Dunn-Walters DK. Human marginal-zone B cells. Immunol Today 1998;19:421–426.
166. Tierens A, Delabie J, Michiels L, et al. Marginal-zone B cells in the human lymph nodes and spleen show somatic hypermutations and display clonal expansion. Blood 1999;93:226–234.
167. MacLennan ICM, Liu YJ. Marginal zone B cells respond to both polysaccharide antigens and protein antigens. Res Immunol 1991;142:436.
168. Martin BG, Kearney JF. Marginal-zone B cells. Nat Rev Immunol. 2002;2:3232–3335.
169. Srivastava B, Quinn WJ, Hazard K, et al. Characterization of marginal zone B cell precursors. J Exp Med 2005;202:1225–1234.
170. Saito T, Chiba S, Ichikawa M, et al. Notch 2 is preferentially expressed in mature B cells and indispensable for marginal zone B lineage development. Immunity 2003;18:675–685.
171. Girkontaite I, Missy K, Sakk V, et al. B cells, regulation of lymphocyte motility and immune responses. Nat Immunol 2001;2:855–862.
172. Cinamar G, Matloublan M, Lesneski MJ, et al. Sphingosine 1-phosphate receptor 1 promotes B cell localization in the splenic marginal zone. Nat Immunol 2004;5:713–720.
173. Dammers PM, Kroese FGM. Recruitment and selection of marginal zone B cells is independent of exogenous antigens. Eur J Immunol 2005;35:2089–2099.
174. Wen L, Brill-Dashoff J, Shinton SA, et al. Evidence of marginal-zone B cell positive selection in spleen. Immunity 2005;23:297–308.
175. Tierens A, Delabie J, Michaels L, et al. Marginal-zone B cells in the human lymph node and spleen show somatic hypermutation and display clonal expansion. Blood 1999;93:226–234.
176. Stein K, Hummel M, Korljuhn P, et al. Monocytoid B cells are distinct from splenic marginal zone cells and commonly derive from unmutated naïve B cells and less frequently from postgerminal center B cells by polyclonal transformation. Blood 1999;94:2800–2808.
177. Willenbrock K, Jungnickel B, Hansmann M-L, et al. Human splenic marginal zone B cells lack expression of activation-induced cytidine deaminase. Eur J Immunol 2005;35:3002–3007.
178. Weller S, Reynaud C-A, Weill J-C. Splenic marginal B cells in humans: where do they mutate their Ig receptor. Eur J Immunol 2005;35:2789–2792.
179. Kanayama N, Cascalho M, Ohmari H. Analysis of marginal zone B cell development in the mouse with limited B cell diversity: role of the antigen receptor signals in the recruitment of B cells to the marginal zone. J Immunol 2005;174:1438–1445.
180. Steiniger B, Timphus E-M, Jacob R, et al. CD27+ B cells in human lymphatic organs—reevaluating the splenic marginal zone. Immunology 2005;116:429–442.
181. Steiniger B, Timpus E-M, Barth PJ. The splenic marginal zone in humans and rodents: an enigmatic compartment and its inhabitants. Histochem Cell Biol 2006;126:641–648.
182. Dierlamm J, Pittaluga S, Wlodarska I, et al. Marginal zone B cell lymphomas of different sites share similar cytogenetic and morphologic features. Blood 1996;87:299–307.
183. Berger F, Felman P, Thieblemont C, et al. Non-MALT marginal B cell lymphomas: a description of clinical presentation and outcome in 124 patients. Blood 2000;95:1950–1956.
184. Tierens A, Delabie J, Pittaluga S, et al. Mutational analysis of the rearranged immunoglobulin heavy chain genes of marginal zone cell lymphomas indicates an origin from different marginal zone B lymphocyte subsets. Blood 1998;91:2381–2386.
185. Ruiz-Ballesteros E, Molleje M, Rodriguez A, et al. Splenic marginal zone lymphoma: proposal of a new diagnostic and prognostic marker identical after tissue and cDNA microarray analysis. Blood 2005;106:1831–1838.
186. Du MQ, Peng HZ, Degan A, et al. Preferential dissemination of B cell gastric mucosa-associated lymphoid tissue (MALT) lymphoma to the splenic marginal zone. Blood 1997;90:4071–4077.
187. Chen LT, Weiss L. Electron microscopy of the red pulp of human spleen. Am J Anat 1972;134:425–457.
188. Weiss L. The red pulp of the spleen: structural basis of blood flow. Clin Haematol 1983;12:375–393.
189. Chen LT, Weiss L. The role of the sinus wall in the passage of erythrocytes through the spleen. Blood 1973;41:529–537.
190. Lockwood CM. Immunological functions of the spleen. Clin Haematol 1983;12:449–465.
191. Bohnsack JF, Brown EJ. The role of the spleen in resistance to infection. Annu Rev Med 1986;37:49–59.
192. Dickerman JD. Bacterial infection and the asplenic host: a review. J Trauma 1976;16:662–668.
193. Singer DB. Postsplenectomy sepsis. Perspect Pediatr Pathol 1973;1:285–311.
194. Hosea SW, Berch CG, Brown EJ, et al. Impaired immune response of splenectomized patients to polyvalent pneumococcal vaccine. Lancet 1981;1:804–807.
195. Drew PA, Kirrow GK, Ferante A, et al. Alterations in immunoglobulin synthesis by peripheral blood mononuclear cells from splenectomized patients with and without splenic regrowth. J Immunol 1984;132:191.
196. Karrer U, Althage A, Odermatt B, et al. On the key role of secondary lymphoid organs in antiviral immune responses studied in antiviral immune responses studied in alymphoplastic (aly/aly) and spleenless (Hox11-/-) mutant mice. J Exp Med 1997;185:2157–2170.
197. Picker LJ, Butcher EC. Physiological and molecular mechanisms of lymphocyte homing. Annu Rev Immunol 1992;10:561–591.
198. Kim CH. The greater chemotactic network for lymphocyte trafficking; chemokines and beyond. Curr Opin Hematol 2005;12:298–304.
199. Sackstein R. The lymphocyte homing receptors: gatekeepers of the multistep paradigm. Curr Opin Hematol 2005;12:444–450.
200. Ebert LM, Schaerli P, Moser B. Chemokine-mediated control of T cell traffic in lymphoid and peripheral tissues. Mol Immunol 2005;42:799–809.
201. Mora JR, van Andrian VH. T-cell homing specificity and plasticity: new concepts and future challenges. Trends Immunol 2006;27:235–243.
202. Streeter PR, Rouse BT, Butcher EC. Immunohistologic and functional characterization of a vascular addressin involved in lymphocyte homing into peripheral lymph nodes. J Cell Biol 1988;107:1853–1862.
203. von Andrian UH, Hasslen SR, Nelson RD, et al. A central role for microvillous receptor presentation in leukocyte adhesion under flow. Cell 1995;82:989–999.
204. Berlin C, Bargatze RF, Campbell JJ, et al. α4 Integrins mediate lymphocyte attachment and rolling under physiologic flow. Cell 1995;80:413–422.
205. Bargatze RF, Jutila MA, Butcher EC. Distinct roles of L-selectin and integrins α4β7 and LFA-1 in lymphocyte homing to Peyer’s patch-HEV in situ: the multistep model confirmed and refined. Immunity 1995;3:99–108.
206. Arbonés ML, Ord DC, Ley K, et al. Lymphocyte homing and leukocyte rolling and migration are impaired in L-selectin-deficient mice. Immunity 1994;1:247–260.
207. Pavalko FM, Walker DM, Graham L, et al. The cytoplasmic domain of L-selectin interacts with cytoskeletal proteins via α-actinin: receptor positioning in microvilli does not require interaction with α-actinin. J Cell Biol 1995;129:1155–1164.
208. Chen S, Springer TA. An automatic braking system that stabilizes leukocyte rolling by an increase in selectin bond number with shear. J Cell Biol 1999;144:185–200.
209. Kanamori A, Kojima N, Uchimura K, et al. Distinct sulfation requirements of selectins disclosed using cells that support rolling mediated by all three selectins under shear flow. L-selectin prefers carbohydrate 6-sulfation to tyrosine sulfation, whereas P-selectin does not. J Biol Chem 2002;277:32578–32586.
210. Vestweber D, Blanks JE. Mechanisms that regulate the function of the selectins and their ligands. Physiol Rev 1999;79:181–213.
211. Lowe JB. Glycosylation in the control of selectin counter-receptor structure and function. Immunol Rev 2002;186:19–36.
212. Hemmerich S, Butcher EC, Rosen SD. Sulfation-dependent recognition of high endothelial venules (HEV)-ligands by L-selectin and MECA 79, an adhesion-blocking monoclonal antibody. J Exp Med 1994;180:2219–2226.
213. Maly P, Thall AD, Petryniak CE, et al. The α-(1,3) fucosyltransferase Fuc-TVII controls leukocyte trafficking through an essential role in L-, E- and P-selectin ligand biosynthesis. Cell 1996;86:643–653.
214. Kawashima H, Petryniak B, Hiraoka N, et al. N-acetylglucosamine-6-O-sulfotransferases 1 and 2 cooperatively control lymphocyte homing through L-selectin ligand biosynthesis in high endothelial venules. Nat Immunol 2005;6:1096–1104.
215. Uchimura K, Ganguet J-M, Singer MS, et al. A major class of L-selectin ligands is eliminated in mice deficient in two sulfotransferases expressed in high endothelial venules. Nat Immunol 2005;6:1105–1113.
216. Warnock RA, Askari S, Butcher EC, et al. Molecular mechanisms of lymphocyte homing to peripheral lymph nodes. J Exp Med 1998;187:205–216.
217. Grabovsky V, Dwir O, Alon R. Endothelial chemokines destabilize L-selectin mediated lymphocyte rolling without inducing selectin shedding. J Biol Chem 2002;227:20640–20650.
218. Kanda H, Tanaka T, Matsumoto M, et al. Endomucin, a sialomucin expressed in high endothelial venules, supports L-selectin-mediated rolling. Intern Immunol 2004;16:1265–1274.
219. Samulowitz V, Kuhn A, Brachtendorf G, et al. Human endomucin-distribution pattern, expression on high endothelial venules, and decoration with the MECA-79 epitope. Am J Pathol 2002;160:1669–1681.
220. Martinez M, Joffraud M, Giraud S, et al. Regulation of PSGL-1 interactions with L-selectin, P-selectin, and E-selectin role of human fucosyltransferase-IV and VII. J Biol Chem 2005;280:5378–5390.
221. Butcher EC, Picker LJ. Lymphocyte homing and homeostasis. Science 1996;272:60–66.
222. Ebnet K, Kaldjian EP, Anderson AO, et al. Orchestrated information transfer underlying leukocyte endothelial interactions. Annu Rev Immunol 1996;14:155–177.
223. Alon R, Feigelson S. From rolling to arrest on blood vessels: leukocyte tap dancing on endothelial integrin ligands and chemokines at sub-second contacts. Sem Immunol 2002;14:93–104.
224. Gunn MD, Tangemann K, Tam C, et al. A chemokine expressed in lymphoid high endothelial venules promotes the adhesion and chemotaxis of naïve T lymphocytes. Proc Natl Acad Sci U S A 1998;95:258–263.
225. Nagira M, Imai T, Yoshida R, et al. A lymphocyte-specific CC chemokine, secondary lymphoid tissue chemokine (SLC), is a highly efficient chemoattractant for B cells and activated T cells. Eur J Immunol 1998;28:1516–1523.
226. Campbell JJ, Butcher EC. Chemokines in tissue-specific and microenvironment-specific lymphocyte homing. Curr Opin Immunol 2000;12:336–341.
227. Campbell JJ, Hendrick J, Zlotnic A, et al. Chemokines and the arrest of lymphocytes under flow conditions. Science 1998;279:381–384.
228. Tangemann K, Gunn MD, Giblin P, et al. A high endothelial cell-derived chemokine induces rapid, efficient, and subset-selective arrest of rolling T lymphocytes on a reconstituted endothelial substrate. J Immunol 1998;161:6330–6337.
229. Ngo VN, Tang HL, Cyster JG. Epstein-Barr virus-induced molecule 1 ligand chemokine is expressed by dendritic cells in lymphoid tissues and strongly attracts naïve T cells and activated B cells. J Exp Med 1998;188:373–386.
230. Backevold ES, Yamanaka T, Palframan RT, et al. The CCR7 ligand ELC (CCL19) is trancytosed in high endothelial venules and mediate T cell recruitment. J Exp Med 2001;193:1105–1111.
231. Kinashi T. Intracellular signalling controlling integrin activation in lymphocytes. Nat Rev Immunol 2005;5:546–559.
232. Nakano H, Mori S, Yonekawa H, et al. A novel mutant gene involved in T-lymphocyte-specific homing into peripheral lymphoid organs on mouse chromosome 4. Blood 1998;91:2886–2895.
233. Fan L, Reilly CR, Luo Y, et al. Cutting edge: ectopic expression of the chemokine TCA4/SLC is sufficient to trigger lymphoid neogenesis. J Immunol 2000;164:3955–3959.
234. Schaerli P, Willimann K, Lang AB, et al. CXC chemokine receptor 5 expression defines follicular homing T cells with B cell helper function. J Exp Med 2000;192:1553–1562.
235. Ansel KM, Ngo VN, Hyman PL, et al. A chemokine-driven positive feedback loop organizes lymphoid follicles. Nature 2000;406:309–314.
236. Luther SA, Lopez T, Bai W, et al. BCL expression in pancreatic islets causes B cell recruitment and lymphotoxin-dependent lymphoid neogenesis. Immunity 2000;12:471–481.
237. Ngo VN, Korner H, Gunn MD, et al. Lymphotoxin alpha/beta and tumor necrosis factor are required for stromal cell expression of homing chemokines in B and T cell areas of the spleen. J Exp Med 1999;189:403–412.
238. Wittchen ES, van Buul JD, Burridge K, et al. Trading spaces: Rap, Rac, and Rho as architects of transendothelial migration. Curr Opin Hematol 2005;12:14–21.
239. Zhang Z, Vuori K, Wang H, et al. Integrin activation by R-ras. Cell 1996;85:61–69.
240. Sotsios Y, Ward SG. Phosphoinositide kinase: a key biochemical signal for cell migration in response to chemokines. Immunol Rev 2000;177:217–235.
241. Ward SG. T lymphocytes on the move: chemokines, PI-3 kinase and beyond. Trends Immunol 2006;27:80–87.
242. Nombela-Arrieta C. Differential requirements for DOCK2 and phosphoinositide-3-kinase γ during T and B lymphocyte homing. Immunity 2004;21:429–441.
243. Dieu MC, Vandervliet B, Vicari A, et al. Selective recruitment of immature and mature dendritic cells by distinct chemokines expressed in different anatomic sites. J Exp Med 1998;188:373–386.
244. Romani N, Ratzinger G, Pfaller K, et al. Migration of dendritic cells into lymphatics. The Langerhans cell example: routes, regulation, and relevance. Int Rev Cytol 2001;207:237–270.
245. Cook DN, Prosser DM, Foster R, et al. CCR6 mediates dendritic cell localization, lymphocyte homeostasis, and immune responses in mucosal tissue. Immunity 2000;12:495–503.
246. Schwarzenberger K, Udey MC. Contact allergens and epidermal pro-inflammatory cytokines modulate Langerhans cell E-cadherin expression in situ. J Invest Dermatol 1996;106:553–558.
247. Moodycliffe AM, Schreedhar V, Ullrich SE, et al. CD40-CD40 ligand interactions in vivo regulate migration of antigen bearing dendritic cells from the skin to draining lymph nodes. J Exp Med 2000;191:2011–2020.
248. Kellerman SA, Hudak S, Oldham ER, et al. The CC chemokine receptor-7 ligands 6C kine and macrophage inflammatory protein-3 β are potent chemoattractants for in vitro- and in vivo-derived dendritic cells. J Immunol 1999;162:3859–3864.
249. Kobayashi Y, Matsumoto M, Kotani M, et al. Possible involvement of matrix metalloproteinase-9 in Langerhans cell migration and maturation. J Immunol 1999;163:5989–5993.
250. Cyster JG. Chemokines and the homing of dendritic cells to the T cell areas of lymphoid organs. J Exp Med 1999;189:447–450.
251. Mackay CR. Migration pathways and immunologic memory among T lymphocytes. Semin Immunol 1992;4:51–58.
252. Mackay CR, Marston WL, Dudler L, et al. Tissue-specific migration pathways by phenotypically distinct subpopulations of memory T cells. Eur J Immunol 1992;22:887–895.
253. Berg EL, Yoshino T, Rott LS, et al. The cutaneous lymphocyte antigen is a skin lymphocyte homing receptor for the vascular lectin endothelial cell-leukocyte adhesion molecule 1. J Exp Med 1991;174:1461–1466.
254. Picker LJ, Kishimoto TK, Smith CW, et al. ELAM-1 is an adhesion molecule for skin-homing T cells. Nature 1991;349:796–799.
255. Cotran RS, Gimbrone MA Jr, Bevilacqua MP, et al. Induction and detection of a human endothelial activation antigen in vivo. J Exp Med 1986;164:661–666.
256. Wayner EA, Gil SG, Murphy GF, et al. Epiligrin, a component of epithelial basement membranes, is an adhesive ligand for α3β1 positive T lymphocytes. J Cell Biol 1993;121:1141–1152.
257. Cerf-Bensussan N, Bègue B, Gagnon J, et al. The human intraepithelial lymphocyte marker HML-1 is an integrin consisting of a β7 subunit associated with a distinctive α chain. Eur J Immunol 1992;22:273–277.
258. Cepek KL, Parker CM, Madara JL, Brenner MB. Integrin αEβ7 mediates adhesion of T lymphocytes to epithelial cells. J Immunol 1993;150:3459–3470.
259. Picker LJ, Terstappen LW, Rott LS, et al. Differential expression of homing-associated adhesion molecules by T cell subsets in man. J Immunol 1990;145:3247–3255.
260. Parker CM, Cepek KL, Russell GJ, et al. A family of β7 integrins on human mucosal lymphocytes. Proc Natl Acad Sci U S A 1992;89:1924–1928.
261. Briskin MJ, McEvoy LM, Butcher EC. MAdCAM-1 has homology to immunoglobulin and mucin-like adhesion receptors and to IgA1. Nature 1993;363:461–464.
262. Tanaka Y, Adams DH, Hubscher S, et al. T-cell adhesion induced by proteoglycan-immobilized cytokine MIP-1β. Nature 1993;361:79–82.
263. Schall TJ, Bacon K, Toy KJ, et al. Selective attraction of monocytes and T lymphocytes of the memory phenotype by cytokine RANTES. Nature 1990;347: 669–671.
264. Miller MD, Krangel MS. Biology and biochemistry of the chemokines: a family of chemotactic and inflammatory cytokines. Crit Rev Immunol 1992;12:17–46.
265. Papayannopoulou T, Craddock C, Nakamoto B, et al. The VLA4/VCAM-1 adhesion pathway defines contrasting mechanisms of lodgment of transplanted murine hemopoietic progenitors between bone marrow and spleen. Proc Natl Acad Sci U S A 1995;92:9647–9651.
266. Zannettino AC, Berndt MC, Butcher C, et al. Primitive human hematopoietic progenitors adhere to P-selectin (CD62P). Blood 1995;85:3466–3477.
267. Puri KD, Finger EB, Gaudernack G, et al. Sialomucin CD34 is the major L-selectin ligand in human tonsil high endothelial venules. J Cell Biol 1995;131:261–270.
268. Kanda T, Yamawaki M, Ariga T, et al. Interleukin 1β up-regulates the expression of sulfoglucuronosyl paragloboside, a ligand for L-selectin, in brain microvascular endothelial cells. Proc Natl Acad Sci U S A 1995;92:7897–7901.
269. Smith ME, Ford WL. The recirculating lymphocyte pool of the rat: a systemic description of the migratory behavior of recirculating lymphocytes. Immunology 1983;49:83–94.
270. Förster R, Schubel A, Breitfeld D, et al. CCR7 coordinates the primary immune response by establishing functional microenvironments in secondary lymphoid organs. Cell 1999;99:23–33.
271. Butcher EC, Williams M, Youngman K, et al. Lymphocyte trafficking and regional immunity. Adv Immunol 1999;72:209–253.
272. Shaniel C, Rolinck AG, Melchers F. Attractions and migrations of lymphoid cells in the organization of humoral immune responses. Adv Immunol 2001;78:111–168.
273. Gerard C, Rollins BJ. Chemokines and disease. Nat Immunol 2001;2:108–115.