Keith M. Skubitz
Three types of granulocytes are readily identified in peripheral blood smears. Neutrophils are so named because of their neutral staining with Wright stain, whereas eosinophils avidly stain with the dye eosin, and basophils have readily identified large dark-staining granules with Wright stain. Neutrophils play a critical role in host defense by phagocytizing and digesting microorganisms, and inappropriate activation of neutrophils may result in damage to normal host tissues. In the resting uninfected host, the production and elimination of neutrophils are balanced, resulting in a fairly constant concentration of neutrophils in peripheral blood. When an infection occurs, chemotactic agents are generated that result in migration of neutrophils to the site of the infection and activation of neutrophil defensive functions. This effect is often associated with an increased production and release of neutrophils from the bone marrow. This chapter reviews the structure, morphology, production, distribution and kinetics, and functions of neutrophils. This chapter is written in several sections, and the first part of each section contains more general information for students, whereas the second part contains greater detail.
![]() Subcellular Structure of Neutrophils
Subcellular Structure of Neutrophils
Mature neutrophils contain several types of granules and other subcellular organelles. To better understand the functions of neutrophils, many studies have been performed to identify and characterize the molecular composition of the various subcellular compartments of neutrophils. Two techniques are commonly used for this purpose: immunoelectron microscopy and subcellular fractionation. Most recent subcellular fractionation studies used the technique of nitrogen cavitation to disrupt the neutrophils. In this technique, cells are equilibrated with nitrogen at high pressure and then released into a pressure of 1 atm. The rapid decrease in extracellular pressure results in disruption of the cells by the formation of nitrogen gas within the cell. The disrupted cells are then fractionated by density-gradient centrifugation (1,2). Many types of subcellular organelles and structures are present in neutrophils, as seen by electron microscopy (EM). Four well-defined types of granules have been defined in neutrophils: primary granules, secondary granules, tertiary granules, and secretory vesicles, although additional heterogeneity of these fractions may exist. Some of the known constituents of these granules are indicated in Table 9.1.
Azurophilic Granules
The azurophilic, or primary, granules are formed during the promyelocytic stage and in general contain many antimicrobial compounds. These granules fuse with phagocytic vesicles, resulting in the delivery of their contents to the ingested organism. Among the azurophilic granule contents is myeloperoxidase (MPO), a protein that catalyzes the production of hypochlorite (OCl-) from chloride and hydrogen peroxide produced by the oxidative burst. MPO constitutes approximately 5% of the dry weight of the neutrophil (18) and imparts the greenish coloration to pus. The human neutrophil defensins, (HNP-1 to -3), a group of cationic proteins that kill a variety of bacteria, fungi, and viruses (19,20,21), also constitute approximately 5% of total neutrophil protein (22). Other components of azurophilic granules include lysozyme, which degrades bacterial peptidoglycans (23), bactericidal permeability-increasing protein, which has antibacterial activity against certain gram-negative bacteria (24,25,26,27,28), azurocidin, which has antibacterial as well as antifungal activity against Candida albicans (29,30), and the serine proteinases elastase, cathepsin G, proteinase 3, esterase N, and others (31,32,33,34). The granule membrane itself contains a large amount of CD66c (35,36) and CD63 antigens (37). Heterogeneity among azurophil granules is likely (3).
Specific Granules
Although some specific (also called secondary) granules, like azurophilic granules, fuse with phagocytic vesicles, it is believed that these granules are largely for release into the extracellular space. The known contents of these granules are also indicated in Table 9.1 and include apolactoferrin, the major specific granule protein, vitamin B12-binding protein, plasminogen activator, and collagenase. Lysozyme and some gelatinase are also present in specific granules. Release of specific granule contents may modify the inflammatory process. For example, collagenase may degrade collagen, thus augmenting movement through collagen and participating in tissue remodeling. Apolactoferrin, by binding iron, may have an antibacterial effect by preventing bacteria from obtaining necessary iron for growth (38). Iron binding by apolactoferrin may also modify hydroxyl radical formation and cell adhesion (39,40,41,42). Although the antimicrobial and proinflammatory defensins are stored in azurophil granules, proHNPs produced at more mature stages of differentiation are not cleaved to the antimicrobial form and are stored in specific granules. ProHNPs are constitutively exocytosed, though their function is unclear (16). Haptoglobin released from specific granules could also inhibit bacterial growth and inflammation (17). Specific granules also contain a number of membrane-bound molecules that are also expressed on the cell surface. This includes CD11, CD18, CD66a, CD66b, NB-1, f-met-leu-phe (FMLP) receptors, C5a receptors, and cytochrome b558. When cells are stimulated, the surface expression of many of these membrane proteins is increased, and some of the up-regulated molecules may be derived from specific granules. The importance of the specific granules in neutrophil function is shown in patients who lack specific granules; these patients are susceptible to repeated skin and respiratory infections and have defective neutrophil chemotaxis and adhesion.
Gelatinase (Tertiary) Granules
Gelatinase, or tertiary, granules, which cosediment with specific granules in some subcellular fractionation techniques, were initially identified as gelatinase-containing granules (43). Like specific granules, tertiary granules also contain many membrane proteins that are up-regulated to the cell surface with stimulation (2). The relative contribution of tertiary granules and specific granules to up-regulation of membrane proteins is not clear.
Secretory Vesicles
Secretory vesicles, which largely distribute in the plasma membrane fraction using subcellular fractionation techniques, have also been described. Complement receptor (CR) 1, recognized by CD35 monoclonal antibodies, has been found exclusively in the light membrane fractions containing secretory vesicles and plasma membranes using subcellular fractionation techniques (12). The observation that CR1 can be readily up-regulated to the neutrophil surface with weak stimulation demonstrates that secretory vesicles provide an intracellular reservoir from which membrane proteins can be recruited to the cell surface. CR3 (HMac-1), recognized by CD11b antibodies and present in both secretory vesicles and specific granules, is also up-regulated to the cell surface with weak stimulation. In contrast to CR1, a more marked up-regulation of CR3 is observed with more potent stimulation, demonstrating that specific granules can also serve as an intracellular reservoir from which membrane proteins can be up-regulated to the cell surface (12). By EM, the secretory vesicles appear as smooth-surfaced vesicles. A defining feature of secretory vesicles is their rapid and complete translocation to the surface membrane with weak stimulation (12). These secretory vesicles also contain alkaline phosphatase, cytochrome b558, and FMLP receptors. Secretory vesicles can be up-regulated to the cell surface in the absence of extracellular calcium, in contrast to specific and gelatinase granules, which require extracellular calcium for release (12). The secretory vesicles appear to be formed by a process of endocytosis and contain albumin.
Plasma Membrane
Many constituents of the neutrophil plasma membrane have been defined. These include membrane channels, adhesive proteins, receptors for various ligands, ion pumps, and ectoenzymes. In the last 15 years, there has been an explosion of information about membrane proteins, identified largely by monoclonal antibodies. Many of these cell-surface molecules probably play a role in regulating the neutrophil response. For example, aminopeptidase N (CD13) can inactivate interleukin (IL)-8, eliminating its chemotactic activity (44), and neutrophil endopeptidase (CD10) can inactivate the chemotactic peptide FMLP (45). Studies with CD66 monoclonal antibodies suggest a role for CD66a, b, c, and d in activating neutrophils (46), and some CD45 antibodies, which recognize a transmembrane protein with tyrosine phosphatase activity in its cytoplasmic domain, inhibit neutrophil chemotaxis (47). Some of the various clusters of differentiation (CD) expressed on neutrophils are shown in Table 9.2. The components of the membrane are not uniformly distributed. Studies have indicated the presence of differentiated domains in the membrane called rafts, which are described in the section “Lipid Rafts.”
|
Table 9.2 Some CD Antigens Expressed on Neutrophils |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Cytoskeletal Matrix
Like many other cells, neutrophils contain a complex cytoskeleton. Alterations in the distribution of cytoskeletal elements may be important in chemotaxis, phagocytosis, and exocytosis. Many protein components of this cytoskeleton have been identified, including actin, actin-binding protein, α-actinin, gelsolin, profilin, myosin, tubulin, and tropomyosin. Actin accounts for approximately 10% of neutrophil protein (48). The reader is referred to other reviews for a more detailed description of the role of the cytoskeleton in neutrophil function (49).
Neutrophil Lipids
Although most studies of neutrophil structure have concentrated on proteins, lipids and carbohydrates also serve important functions. Lipids account for approximately 5% of neutrophils by weight (50,51), of which approximately 35% is phospholipid (52). Phosphatidylcholine and phosphatidylethanolamine account for approximately 75% of the phospholipid in intact neutrophils (52). Subcellular distribution studies reveal that plasma membranes and secretory vesicles contain approximately 50% of the cellular phospholipid. Most of the phosphatidylinositol and phosphatidylcholine are present in plasma membrane and secretory granules, whereas a large part of the phosphatidylethanolamine is found in the specific granules (52). Among the phospholipids, the phosphoinositides are important as sources of inositol 1,4,5-triphosphate (IP3) (a signal-transduction molecule that results in calcium release) and diacylglycerol (which activates protein kinase C [PKC]). The occurrence of arachidonic acid in phospholipids, especially phosphatidylcholine, is important as a precursor for the production of leukotrienes, prostaglandins, thromboxanes, and lipoxins (53,54). Cholesterol and triglycerides constitute most of the nonphospholipid neutrophil lipid. Glycolipids, which include both neutral glycosphingolipids and gangliosides, constitute the remaining neutrophil lipids. The study of glycolipids is complex, and the glycolipid composition of neutrophils is not well understood. Glycolipids are important because their carbohydrate components contain many neutrophil differentiation antigens with a multitude of potential functions. The major neutrophil glycolipid is lactosylceramide (LacCer: Galβ1 →4Glcβ1 → 1Cer) (55,56,57) and is recognized by CDw17 monoclonal antibodies. Interestingly, the surface expression of LacCer decreases after neutrophil stimulation. More than 75% of neutrophil LacCer is found in intracellular granules (56,58). It has been hypothesized that most LacCer in the granule membranes is found in the outer leaflet and may contribute to the ability of these membranes to form the highly convex surfaces necessary to form these submicrometer particles (54). Approximately 75% of the five major glycosphingolipids are located intracellularly (58). Studies of the subcellular distribution of glycosphingolipids in neutrophils have found no differences among the plasma membrane, primary granules, or secondary granules in the relative amounts of these five glycosphingolipids (58).
Lipid Rafts
Studies have demonstrated the existence of large noncovalent detergent-resistant complexes in cell extracts that contain important signaling molecules, including protein kinases and many glycosyl-phosphatidylinositol–linked membrane proteins capable of transmitting signals (59,60,61,62). These complexes have been termed lipid rafts or large detergent-resistant complexes. It is postulated that these complexes or rafts reflect the existence of specific membrane microdomains that have a particular lipid composition, and that these clusters of molecules may be important in transmembrane signaling by proteins in the complex. Rafts or detergent-resistant complexes have been observed in neutrophils (63; Draber P, Draberova L, Skubitz K, personal communication, 2001). There is evidence that in neutrophils, proteins may enter these rafts when they are translocated to the cell surface. For example, it appears that CD63 and CD11b/CD18 are not present in detergent-resistant complexes when they are intracellular, but they enter such complexes after translocation to the cell surface (63).
Cytoplasmic Lipid Bodies
Cytoplasmic lipid bodies, non–membrane-bound cytoplasmic inclusions, have been described in neutrophils (57). In inflammation, the number of cytoplasmic lipid bodies in neutrophils increases (65). These lipid bodies may provide nonmembrane stores of esterified arachidonate. In addition, some signaling proteins, including phosphatidylinositide-3-kinase, are localized to these lipid bodies (66), although the exact role of these lipid bodies in cell function is unclear (66,67).
Cytosol
Although neutrophil cytoplasm contains many components common to all cells, it is interesting to note that approximately 45% of neutrophil cytosolic protein appears to be attributable to migration inhibitory factor–related proteins (MRPs), MRP-8 and MRP-14 (68). MRP-8 and MRP-14 are members of the S100 family of calcium-binding proteins and form homo- and heterodimers. MRP-14 has been variously called p14, L1 heavy chain, and calgranulin b, and MRP-8 is also known as p8, L1 light chain, calgranulin a, and cystic fibrosis antigen(69,70,71,72,73). Although the role of these proteins in neutrophil function is unclear, the quantity of MRP-8 and MRP-14 associated with neutrophil plasma membranes has been reported to increase after stimulation (68). MRP-8/MRP-14 are also present in secondary and/or tertiary granules and are released when neutrophils are stimulated (73). MRP-14 can inhibit macrophage activation (74). Annexin I or lipocortin I comprises approximately 3% of cytosolic protein (75). Annexin I is partially regulated by glucocorticoids and appears to be a mediator of the anti-inflammatory effects of glucocorticoids (75).
One other notable cytoplasmic constituent is glycogen. Because neutrophils are sometimes required to function in hypoxic conditions, as in an abscess, they are very capable of obtaining energy by glycolysis. The presence of large intracellular glycogen stores gives them the additional ability to function in areas of low extracellular glucose.
Nucleus
In the past, it has been felt that neutrophils, as end-stage cells, undergo little RNA or protein synthesis. More recently, it has been demonstrated that mature neutrophils can synthesize both RNA and protein (76,77,78).
![]() Morphology of Neutrophils and Precursors
Morphology of Neutrophils and Precursors
Evidence for the replenishment of marrow and blood cells from a stem cell compartment is described in Chapter 8 (73). Neither multipotent hematopoietic stem cells nor more committed progenitors are readily identified morphologically by traditional methods, and reliable EM criteria for distinguishing myeloblasts from monoblasts or lymphoblasts are also lacking (79,80,81,82,83), although pronormoblasts often can be differentiated by the presence of ferritin on the cell surface or in coated vesicles (79). Only the more mature forms of each hematopoietic cell series can be reliably distinguished from one another. In the following pages, the cells identifiable as neutrophils and their precursors are described; other aspects of the eosinophil and basophil systems are considered in Chapters 10 and 11.
Neutrophilic, eosinophilic, and basophilic granulocytes are thought to follow similar patterns of proliferation, differentiation, maturation, and storage in the bone marrow and delivery to the blood. The details of these processes are best documented for neutrophils. In the first three morphologic stages, the myeloblast, promyelocyte, and myelocyte cells are capable of replication, as shown by their uptake of tritiated thymidine (3H-TdR) and the presence of mitoses; in later stages, cells cannot divide but continue to differentiate. The morphologic boundaries of each cell compartment were defined many years ago and were based on criteria such as cell size, ratio of size of nucleus to cytoplasm, fineness of nuclear chromatin, nuclear shape, the presence or absence of nucleoli, the presence and type of cytoplasmic granules, and the cytoplasmic color of stained cells (Table 9.3).
Because changes in nuclear chromatin and cell size occur during each cell replication cycle and the formation of granules and other cytoplasmic changes occur gradually during the stages of cell development, morphologic definitions are necessarily arbitrary and do not always conform to significant biochemical or physiologic changes. Classifying a cell in one category or another often is difficult because it is actually in transition between the two. Nevertheless, it is useful to separate the cell lines into morphologic compartments and to define normal limits of cell distribution therein, because gross changes from these patterns indicate disease.
![]() Development of Neutrophils and Their Precursors
Development of Neutrophils and Their Precursors
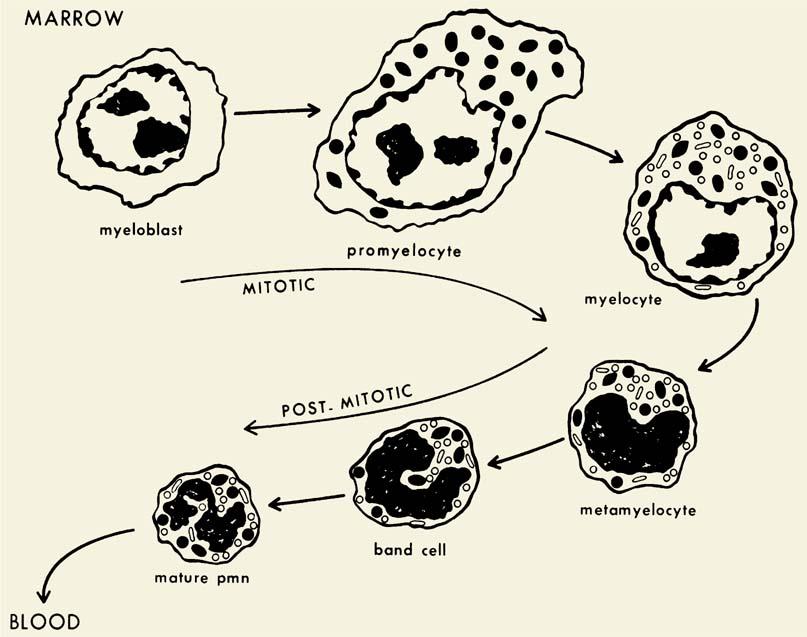
Neutrophil development is summarized in Figure 9.1.
Cell division is limited to myeloblasts, promyelocytes, and myelocytes, with later developmental stages undergoing differentiation but no further cell division. The myeloblast contains few granules and is derived from more primitive cells as described in Chapter 5. As the cell differentiates to the promyelocyte stage, development of primary or azurophil granule formation becomes evident. This granule contains MPO, an enzyme whose activity is a classic marker of myeloid differentiation. Azurophil granule production ceases at the end of the promyelocyte stage, coincident with the loss of peroxidase activity from the rough endoplasmic reticulum. Secondary granule, or specific granule, formation begins as the neutrophil enters the myelocyte stage. The peroxidase-negative specific granules are smaller (approximately 200-nm diameter) than the azurophil granules (approximately 500-nm diameter) and are near the limit of resolution by light microscopy. The specific granules impart a pinkish ground-glass background color to neutrophils in Wright-stained smears. Because azurophil granule formation ceases in the promyelocyte stage and the subsequent myelocyte form is still capable of cell division, the density of azurophil granules is lower in differentiation stages past the promyelocyte. The result is that mature neutrophils contain approximately two specific granules for every azurophil granule. With maturation, the azurophil granules, which generate reddish-purple staining in the promyelocytes, lose this metachromasia as they leave the myelocyte stage. This alteration in staining properties is thought to be caused by an increase in acid mucosubstances, which complex with basic proteins already present in the azurophil granules (84). Thus, in the mature neutrophil, the azurophil granules appear as light blue-violet granules on Wright-stained smears. The azurophil granules are readily demonstrated by peroxidase staining with light microscopy.
Data concerning antigenic differences between granulocytes and monocytes and their stages of maturation have largely been developed using monoclonal antibodies. Such monoclonal antibodies have been analyzed in a series of international workshops in which antibodies are grouped into clusters of differentiation (CD). Some of the CD antigens expressed on neutrophils are shown in Table 9.2. Immunogold and enzyme-linked immunologic methods (85) permit simultaneous morphologic and immunologic examination of individual cells.
Myeloblast
The word myeloblast describes an immature cell, typically found in the bone marrow and not in the blood. This cell can divide and give rise to promyelocytes, which in turn give rise to myelocytes. On the basis of the findings from marrow culture and transplant studies, the neutrophil and macrophage lines share a common stem cell, colony-forming unit granulocyte-monocyte (CFU-GM) (86,87,88,89).
The myeloblast (Fig. 9.2) has a large nucleus, is round or slightly oval, and has a small amount of cytoplasm. In preparations treated with Wright stain (Table 9.3), the nuclear membrane is smooth and even in outline and is exceedingly thin, with no condensation of chromatin near its inner surface, as noted in lymphoblasts. The chromatin shows an even, diffuse distribution with no aggregation into larger masses, although some condensation may be noted about the nucleoli. The chromatin may appear in the form of fine strands, thus giving the nucleus a sievelike appearance; alternatively, it may have the form of fine dustlike granules, producing a uniform stippled effect. Generally, the myeloblast contains two to five pale, sky-blue nucleoli. The cytoplasm is basophilic (blue), and usually, although not invariably, no clear zone is evident about the nucleus. Sometimes, the cytoplasm is reticular, spongy, or foamy. By definition, no granules are present in the cytoplasm. Leukemic myeloblasts that contain no perceptible granules often are identified by special stains that demonstrate the presence of MPO or esterase, thus providing early evidence of differentiation (90).
|
Table 9.3 Morphologic Characteristics of Leukocytes (Wright Stain) |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
EM reveals similar findings (91,92). The nuclear membrane is thin and indistinct, with minimal or no chromatin condensation. The numerous particles of ribonucleoprotein in the cytoplasm produce deep blue basophilia in stained preparations. Mitochondria are abundant but small, and the endoplasmic reticulum is flat and appears infrequently. The Golgi apparatus is indistinct, and no cytoplasmic granules are present. EM studies of myeloblasts show peroxidase activity in the rough endoplasmic reticulum and Golgi.
Some authors classify what may be slightly more mature cells with several rather large, angular, irregular, and dark-staining azurophilic cytoplasmic granules as myeloblasts. A simpler approach, however, is to include such forms in the promyelocyte stage, thus making the separation between the two cell types clear-cut. The EM classification of myeloid cells, which is based primarily on stages of granule formation, also places cells with beginning granule formation in the promyelocyte category (83,93).
|
|
|
Figure 9.1. Appearance of granules during neutrophil maturation. Myeloblasts are undifferentiated cells with a large oval nucleus, large nucleoli, and cytoplasm lacking granules. They originate from a precursor pool of stem cells. Subsequently, there are two stages—the promyelocyte and the myelocyte—each of which produces a distinct type of secretory granule: azurophils (dark granules) are produced only during the promyelocyte stage; specific granules (light granules) are produced during the myelocyte stage. The metamyelocyte and band forms are nonproliferating stages that develop into the mature polymorphonuclear neutrophil characterized by a multilobulated nucleus and cytoplasm containing primarily glycogen and granules. Both nonspecific azurophilic granules and specific granules persist throughout these later stages. (Modified from Bainton DF, et al. The development of neutrophilic PMN leukocytes in human bone marrow: origin and content of azurophil and specific granules. J Exp Med 1971;134:907.) |
|
|
|
Figure 9.2. Myeloblast (×1,000, Wright stain). |
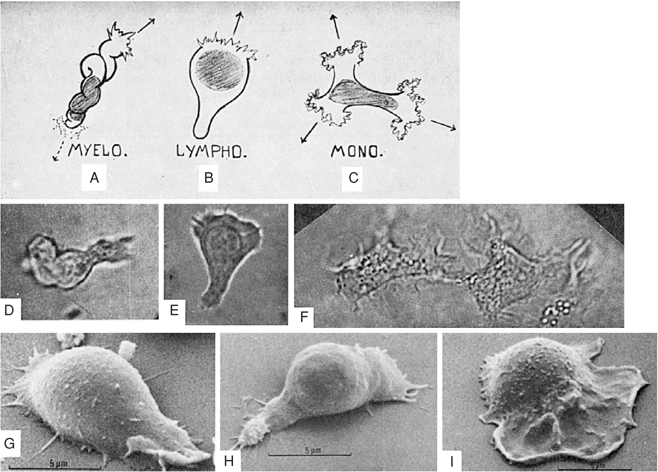
In wet films, myeloblasts appear immobile, with thin, tenacious borders. The cytoplasm is hazy and usually contains no stainable substance other than mitochondria, which are diffusely scattered throughout the cytoplasm and stain brilliant blue-green with Janus green. The failure of myeloblasts to move in supravital preparations is probably related to the nature of the preparation itself rather than to the cells’ immobility. In motion-picture studies of hanging-drop preparations (Fig. 9.3), myeloblasts manifest a characteristic snaillike movement (94,95).
Because they are in the process of growth and division, myeloblasts vary considerably in size from 10 to 20 μm in diameter. Particularly in patients with acute leukemia, the nucleus may show several wide and deep indentations, suggesting lobulation. Such myeloblasts [see Rieder cells (96)] suggest more rapid maturation on the part of the nucleus than of the cytoplasm (asynchronism of Di Guglielmo). Also in association with leukemia, Auer bodies are evident in the cytoplasm of cells that otherwise look like myeloblasts (Fig. 9.4).
Rieder cells, in which the nucleus is polymorphous or highly differentiated yet the cytoplasm is immature, are not a specific cell type but probably represent asynchrony of nuclear and cytoplasmic differentiation in monocytes, lymphocytes, myeloblasts, leukoblasts, or reticuloendothelial monocytoid cells (97).
|
|
|
Figure 9.3. Diagrams (A, B, C), photographs (D, E, F), and scanning electron micrographs (G, H, I) of living, moving, unstained blasts. The photographs are of cells enlarged (×1,000) from negatives of motion picture films of tissue cultures. A: Wormlike shape of myeloblast (myelo.) in motion. B: Hand-mirror shape of lymphoblast (lympho.) in motion. C: Shape of monocyte (mono.) (histiocyte) in motion. D: Myeloblast from the blood of a patient with acute myeloblastic leukemia. E: Lymphoblast from the blood of a patient with acute lymphoblastic leukemia. F: Two monocytes from normal blood. G: Human leukemic myeloblast. H: Human leukemic lymphoblast. I: Human leukemic monoblast. (Photographs used with permission from Rich AR, et al. The differentiation of myeloblasts from lymphoblasts by their manner of locomotion. Bull Johns Hopkins Hosp 1939;65:291; and electron micrographs used with permission from Senda N. The movement of leucocytes. J Clin Electron Microsc 1974;7:3.) |
|
|
|
Figure 9.4. A, B: Pseudo–Pelger-Huët cells, the latter from the blood of a patient with acute myeloblastic leukemia (×1,000, Wright stain). |
Differentiation from Lymphoblasts and Other Blasts
With the usual Romanowsky stains and light microscopy, distinguishing among the leukoblasts (myeloblasts, lymphoblasts, and monoblasts) and even pronormoblasts is extremely difficult if cells showing beginning maturational changes (granule formation or hemoglobin synthesis) are excluded from the blast category (83,91). In the lymphoblast, the nuclear membrane is more dense than that of the myeloblast, and the chromatin is more coarse and may show some aggregation. Figure 9.5 shows representative acute lymphoblastic leukemia cells (Fig. 9.5A,B) and acute myeloid leukemia cells (Fig. 9.5C,D). Lymphoblasts generally have only one or two nucleoli, and their membrane usually is distinct. The mitochondria are short and plumper than those of myeloblasts and often assume a position close to the nucleus. The monoblast is described as showing characteristics similar to those of the mature monocyte, such as fine chromatin, pale nucleus, and ground-glass cytoplasm with a fine, irregular border. In many instances, the identification of blast cells is greatly aided by the company they keep (the more mature and more easily recognized cells about them in sections or in the same blood smear). With the myeloblast, the demonstration of associated promyelocytes, which show azure granulation in Wright or similar stains, is presumptive evidence for this cell’s identification. Special stains to detect enzymes, such as peroxidase or esterase, in the blast cytoplasm before lysosomal granules appear may at times be useful in identification, especially in the classification of leukemia (90,98).
|
|
|
Figure 9.5. A: L1 lymphoblastic leukemia, blood. B: L2 lymphoblastic leukemia, blood. C: Acute leukemia, M1 blood (×1,500). D: Acute leukemia, M1 marrow (×1,500). |
Neutrophil Promyelocytes and Myelocytes
The developmental stages in the granulocyte series and some of their morphologic variations are shown in Figure 9.1. In the past, the several stages beyond the myeloblast were differentiated primarily on the basis of the number and type of granules. Now, EM histochemical and biochemical findings demonstrate that the azurophilic or primary granules first appear at the promyelocyte stage and can be identified on fine structural study as characteristic of the neutrophil, eosinophil, or basophil series (83,91,93,99,100). They do not transform into specific granules but persist throughout the remainder of the maturation sequence and are seen in all subsequent stages, including the polymorphonuclear forms (Fig. 9.1) (93,100,101).
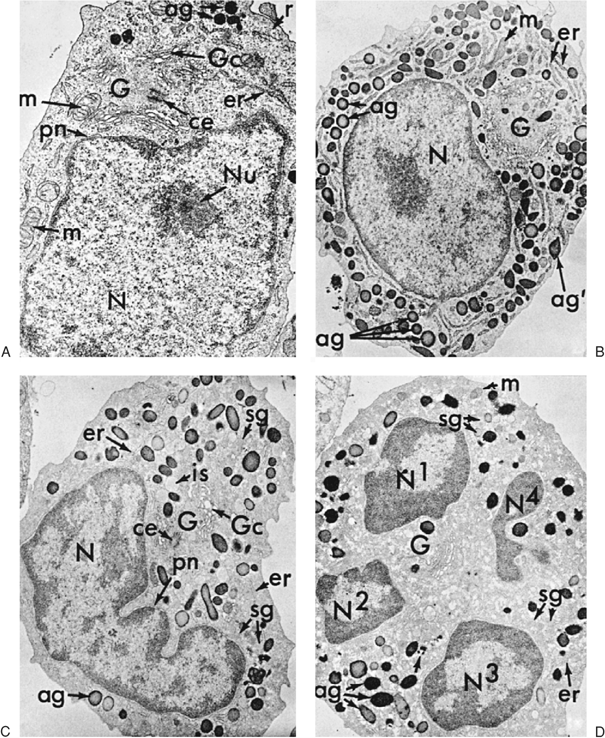
The neutrophil promyelocyte is somewhat larger on average than the myeloblast. In both light and EM preparations, it has a round or oval nucleus in which the nuclear chromatin is diffusely distributed, as in the myeloblast; in later stages, slight chromatin condensation is discerned around the nuclear membrane. Nucleoli are present, but as the cell develops, they become less prominent. Compared with the myeloblast, the endoplasmic reticulum in EM preparations is more prominent and takes on a dilated, vesicular appearance. The azurophilic, primary granules appear and accumulate in increasing numbers during this stage, but the specific or secondary granules are not yet present (Fig. 9.6) (83,92,93,99,101). In early promyelocytes, the few granules present may be difficult to see by using light microscopy; they often lie over the nucleus and are evident only on examination at several focal planes.
Like the myeloblast, the promyelocyte is immobile in flat slide and cover glass preparations; only in the last stage is slight locomotion evident. Even then, the streaming of granules so characteristic of mature granulocytes is lacking (102). For this reason, the cytoplasm has been thought to be in the form of a gel; the increased resistance of the cytoplasm of immature myeloid cells to changes in shape has been proposed as a factor in retention of these cells in the bone marrow (103,104). In hanging-drop preparations, however, promyelocytes are actively mobile.
The neutrophilic myelocyte may be defined as the stage in which specific (secondary) granules appear in the cytoplasm and the cell consequently can be identified as belonging to the neutrophilic series when stained and observed to have a pinkish ground-glass background color with the light microscope. As mentioned previously, earlier identification of a cell that will become neutrophilic can be made by EM examination of the azurophilic, or primary, granules (91). The nucleus of the neutrophilic myelocyte usually is eccentric and round or oval; one side may appear flattened. The nuclear chromatin is somewhat coarse, and nucleoli are small and often not visible, although they are seen clearly with the electron microscope (99). Primary granules persist in myelocytes, but formation of new primary granules is limited to the promyelocyte, and each succeeding cell division leads to a decrease in their number in the daughter population (Fig. 9.1) (93,101,102). The secondary granules of the neutrophil series are smaller than the primary granules; in the rabbit, cat, and human, they are formed in increasing numbers on the convex surface and lateral borders of the somewhat less prominent Golgi apparatus (93,99,101). The amount of granular endoplasmic reticulum is lower in the myelocyte than in earlier forms, so the cytoplasmic basophilia decreases and disappears. The mitochondria remain small and are few.
Neutrophil Granule Development
Studies in the rabbit, cat, and human suggest that the primary granules are packaged and released from the inner, concave surface of the Golgi apparatus (Fig. 9.7A)—in contrast to the specific or secondary granules of the myelocyte and later granulocyte stages that appear to be formed and released from the outer, convex surface (93,99,100,101). Studies of membranes from rabbit azurophil and specific granules, although demonstrating similar ultrastructure, have shown them to be distinct and different in cholesterol–phospholipid ratios and protein components (105). This finding not only appears to confirm different sites of granule formation, it also may provide a basis for differences in interaction with endocytic vacuoles. In the mature neutrophil, a ratio of secondary to primary granules of approximately 9:1 is seen in the rabbit (106) and 2 or 3:1 in humans (93,107).
|
|
|
Figure 9.6. Early and late promyelocytes, a myelocyte, and a polymorphonuclear neutrophil (PMN) viewed by electron microscopy. (Courtesy of Bainton D, University of California, San Francisco.) A: Early neutrophilic promyelocyte (reacted for peroxidase, 10,500). The nucleus (N) with its prominent nucleolus (Nu) occupies the bulk of this immature cell. The surrounding cytoplasm contains a few azurophil granules (ag), a large Golgi complex (G), Golgi cisternae (Gc), several mitochondria (m), scanty rough endoplasmic reticulum (er), and many free polysomes (r). A centriole (ce) is present in the Golgi region. All of the azurophil granules (ag) appear dense because they are strongly reactive for peroxidase. The secretory apparatus, i.e., the perinuclear cisterna (pn), rough endoplasmic reticulum (er), and Golgi cisternae (Gc), are also reactive, although less so than the granules. Specimen was fixed in glutaraldehyde for 16 hours at 4°C, incubated in the peroxidase medium of Graham and Karnovsky for 1 hour at 22°C, postfixed in osmium tetroxide, treated in block with uranyl acetate, dehydrated in ethanol, infiltrated with propylene oxide, and embedded in Araldite. Section stained for 1 minute with lead citrate. B:Late neutrophilic promyelocyte (reacted for peroxidase, 7,000). This cell is the largest (15 μm) of the neutrophilic series. It has a sizable, slightly indented nucleus (N), a prominent Golgi region (G), and cytoplasm packed with peroxidase- positive azurophil granules (ag). Note the two general shapes of the azurophil granules: spherical (ag) and ellipsoid (ag′). Most are spherical, with a homogeneous matrix, but a few ellipsoid forms containing crystalloids also are present. Many of the spherical forms (ag) have a dense periphery and a lighter core, presumably because of incomplete penetration of substrate into the compact centers of mature granules. Peroxidase reaction product is visible (under higher magnification) in less concentrated form within all compartments of the secretory apparatus (endoplasmic reticulum, perinuclear cisterna, and Golgi cisternae). No reaction product is seen in the cytoplasmic matrix, mitochondria, or nucleus. Specimen was fixed in glutaraldehyde for 10 minutes at 4°C and subsequently processed exactly as was the specimen in A. C: Neutrophilic myelocyte (reacted for peroxidase, 9,000). At this stage, the cell is smaller (10 μm) than the promyelocyte, the nucleus is more indented, and the cytoplasm contains two different types of granules: large, peroxidase-positive azurophils (ag) and the generally smaller, specific granules (sg), which do not stain for peroxidase. A number of immature specific granules (is), which are larger, less compact, and more irregular in contour than mature granules, are seen in the Golgi region (G). Note that peroxidase reaction product is present only in azurophil granules and is not seen in the rough endoplasmic reticulum (er), perinuclear cisterna (pn), and Golgi cisternae (Gc), in keeping with the fact that azurophil production has ceased, and only peroxidase-negative specific granules are produced during the myelocyte stage. D: Mature PMN (reacted for peroxidase, 10,500). The cytoplasm is filled with granules; the smaller peroxidase-negative specific granules (sg) are more numerous, the azurophils (ag) having been reduced in number by cell divisions after the promyelocyte stage. Some small, irregularly shaped azurophil granule variants are also present (unlabeled arrow). The nucleus is condensed and lobulated (N1–N4), the Golgi region (G) is small and lacks forming granules, the endoplasmic reticulum (er) is scanty, and mitochondria (m) are few. Note that the cytoplasm of this cell has a rather ragged, moth-eaten appearance because the glycogen, which is normally present, has been extracted in this preparation by staining in block with uranyl acetate. |
|
|
|
Figure 9.7. Granule formation in neutrophil precursors viewed by electron microscopy. (Courtesy of Bainton D, University of California, San Francisco.) A: Golgi region of a neutrophilic promyelocyte reacted for peroxidase (×40,000). At this stage, the peroxidase reaction product is present within the rough endoplasmic reticulum (er), the clusters of smooth vesicles (ve) at the periphery of the Golgi cisternae (Gc), in the Golgi cisternae, and in the immature (ia) and mature (ag) azurophilic granules. The immature granules are larger and less compact than the uniformly dense mature granules. B: Golgi region of a neutrophilic myelocyte reacted for peroxidase (×40,000). Peroxidase-reactive material is seen in the primary or azurophilic granules (ag) but not in the specific (secondary) granules (sg). At this stage (myelocyte), no peroxidase reaction product is seen in the endoplasmic reticulum, Golgi cisternae (Gc), or newly formed, immature specific granules (is). The stacked Golgi cisternae are oriented around the centriole (ce), and the outer cisternae (unlabeled arrow) contain material of intermediate density that is similar to the content of the specific granules, suggesting that the specific granules arise from the convex face of the Golgi complex as in the rabbit. pn, perinuclear cisternae. |
The mature primary granules of human neutrophils usually contain central crystalloids when lightly stained (100). They apparently bind neutral red dye and thus are seen easily as neutral red bodies in supravital preparations (108). These membrane-bound lysosomes contain enzymes and other substances (Table 9.1) (83,93,109,110). Acid phosphatase activity varies considerably, as reported in different studies and in different species (99,100,111,102). This may be because of inadequacies of the histochemical assays or perhaps is related to species variations (83). Peroxidase has been associated with primary granules by histochemical, cytochemical, and biochemical methods and is considered a marker enzyme for primary granules in mammals (83,113). Sulfated mucosubstance presumably accounts for the azurophilic staining of the primary granules; the uptake of radiosulfate by early-stage neutrophils may be the result of incorporation into this substance (114,115).
The secondary granules of the neutrophil were thought in the past to be characterized by their content of alkaline phosphatase and lack of acid phosphatase (83,93,116); they lack peroxidase and sulfated mucosaccharide or contain a minimal amount (83). Alkaline phosphatase was found in blood and exudate neutrophils by many workers, but considerable variation exists between species (83,99,109,111) and in certain pathologic states (117). Results of histochemical (83,93,111,118) and biochemical (116,119) studies suggested that this enzyme was located in the secondary granules of neutrophils in humans and rabbits, perhaps bound to the inner membrane (107,120). However, subsequent studies demonstrated that alkaline phosphatase is localized in a previously unrecognized organelle, the secretory vesicle (Table 9.1) (121). Lysozyme is present in human neutrophil secondary granules (122), as well as in primary granules; approximately two thirds of this antibacterial basic protein is in the secondary granule (89,119). The standard marker enzymes for specific granules are lactoferrin (113) and B12-binding protein (123).
A third type of granule, the tertiary granule, also known as the gelatinase granule (Table 9.1) (100,124), is synthesized mainly during the band and segmented neutrophil stages (100,125,126).
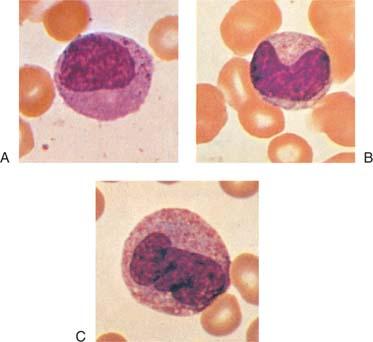
Neutrophil Metamyelocytes
The metamyelocyte is characterized by a clearly indented or horseshoe-shaped nucleus without nucleoli (even by EM examination), and the nuclear chromatin is moderately dense, with considerable clumping evident along the nuclear membrane. The cytoplasm is filled with primary, secondary, and tertiary (88,114) granules, but the secondary granules predominate. The endoplasmic reticulum is sparse, as are polysomes, thus signifying the virtual completion of protein synthesis.
The boundary between the myelocyte and metamyelocyte compartments is best defined physiologically by the fact that myelocytes synthesize DNA, take up 3H-TdR into their nuclear chromatin, divide, and are actively involved in protein synthesis, as evidenced by the presence of nucleoli, abundant endoplasmic reticulum, and polysomes. Before such techniques became available, differentiation between myelocytes and metamyelocytes was defined mainly in terms of nuclear shape. This characteristic now is recognized as a poor criterion because it has been shown in time-lapse microcinematographic studies of human neutrophils that myelocyte nuclei may assume a markedly indented shape and may subsequently revert to an oval configuration and enter mitosis (91). Consequently, in classifying cells at this stage, the observer should pay particular attention to evidence in the nucleus and cytoplasm that protein synthesis has decreased or stopped. This determination is made on the basis of the fact that the nuclear chromatin is coarse and clumped and that the cytoplasm is faint pink and is essentially the color of the mature cell in stained preparations. These features also are helpful in differentiating metamyelocytes (Fig. 9.8A,B) from monocytes (Fig. 9.8C; see also Fig. 12.3) because in monocytes, nuclear chromatin remains fine, and evidence of protein synthesis persists. Ameboid movement is apparent in metamyelocytes, even in cover glass slide preparations, and it is at this stage that directional migration can regularly be demonstrated (127).
|
|
|
Figure 9.8. A: Late myelocyte or early metamyelocyte. B: Metamyelocyte. C: Monocyte (×1,000, Wright stain). |
Band Neutrophils
The band stage is characterized by further condensation of nuclear chromatin and transformation of nuclear shapes into sausage or band configurations that have approximately uniform diameters throughout their length (Fig. 9.1). Subsequently, one or more constrictions begin to develop and progress until the nucleus is divided into two or more lobes connected by filamentous strands of heterochromatin, the polymorphonuclear stage. A difference of opinion exists concerning the differentiation of the juvenile (band) and polymorphonuclear stages. Some workers require a clearly visible filamentous strand between lobes (Fig. 9.9A,B) before classifying a cell as a polymorphonuclear form; anything less clear-cut, whether because of overlapping of nuclear lobes or incomplete constriction, is classified as a band form (128,129). Other investigators regard a constriction greater than one half or two thirds of the nuclear breadth as adequate evidence of lobulation and classify such cells as polymorphonuclear (130,131) or use slightly different criteria (132). Still others avoid the issue entirely. Because no clear difference has been shown between band and segmented stages other than nuclear shape and a slightly earlier appearance of 3H-TdR in the band forms, the distinction becomes arbitrary. However, a clear and easily recognizable separation is needed if one wishes to count nuclear lobes for diagnostic purposes, as in the early detection of folic acid deficiency (133) or in assessing marrow release of young forms into the blood (134). For such purposes, we have chosen the clear separation of nuclear lobes as the criterion for inclusion in the polymorphonuclear category (129). Cells without this complete formation of distinct lobes (usually connected by a filamentous strand) are classified as band forms.
|
|
|
Figure 9.9. A, B: Polymorphonuclear neutrophils. |
Pelger-Huët Anomaly
Pelger-Huët anomaly is a benign anomaly of leukocytes and is inherited as a non–sex-linked, dominant trait. It is characterized by distinctive shapes of the nuclei of leukocytes, a reduced number of nuclear segments (best seen in the neutrophils), and coarseness of the chromatin of the nuclei of neutrophils, lymphocytes, and monocytes. The nuclei appear rodlike, dumbbell-shaped, peanut-shaped, and spectaclelike (“pince-nez”) with smooth, round, or oval individual lobes (Fig. 9.4), contrasted with the irregular lobes seen in normal neutrophils. Pelger-Huët cells appear to be normal functionally. Pseudo– or acquired Pelger-Huët anomaly may also be seen, in which cells with morphologic changes, such as described above, have been observed occasionally in association with myxedema, acute enteritis, agranulocytosis, multiple myeloma, malaria, leukemoid reactions secondary to metastases to the bone marrow, drug sensitivity, or chronic lymphocytic leukemia. More commonly, pseudo–Pelger-Huët cells are seen in patients with myeloid leukemia or myeloid metaplasia. Pelger-Huët anomaly is discussed further in Chapter 64.
Polymorphonuclear Neutrophils
In the polymorphonuclear stage, the nucleus in Wright-stained preparations is a deep purplish color and contains course, condensed chromatin. The lobes are joined by thin filaments of chromatin, although the filaments may not be easily visible if the lobes are partially superimposed. Careful examination by focusing through several planes may facilitate identification. The cytoplasm is faint pink and contains fine, specific granules that sometimes give only a ground-glass appearance. The azurophilic or primary granules have usually lost their dark-staining characteristics by this stage but can be seen with EM. With this technique, the granules exhibit considerable variation in density, presumably a reflection of variation in enzyme content, and they maintain a minimum distance of 100 nm from the cell membrane (135). Large masses of glycogen become evident for the first time in mature neutrophils; this finding may contribute to their capacity for anaerobic metabolism.
The mechanism and purpose of nuclear lobulation are the subject of much speculation. Perhaps it enhances cell deformability and movement through vessel walls and into sites of inflammation, or perhaps nuclear segmentation results from nucleolar emptying and has no function (136). In studies of the nuclear protein matrix at different stages of neutrophil maturation, researchers found no significant changes other than collapse and an increased binding of DNA as the cell matures (137). Thus, the mechanism and purpose of nuclear segmentation remain unclear.
Arneth believed that nuclear lobulation continues as the cell ages and that granulocytes with three or four lobes are more mature than those with only two (138). This statement may be true in the sense that once nuclear constriction begins, the completion of lobulation may continue for some time; during this interval, the cell may be delivered to the blood and thus completion of lobulation may occur in the blood (or in cultures). However, the number of lobes a neutrophil develops appears to be determined in the band stage (or earlier), and the time of appearance of neutrophils in the blood after pulse labeling with 3H-TdR is unrelated to the number of nuclear lobes (139).
In wet films, marked ameboid activity of polymorphonuclear neutrophils at physiologic temperatures is characteristic (140). In supravitally stained films, the (specific) granules appear yellowish pink and are refractile; in addition, occasional larger, nonrefractile, deep red vacuoles may be seen.
“Senile” polymorphonuclear leukocytes that are no longer motile and fail to take up the neutral red stain have been identified in in vitro preparations (141). They are seen in small numbers in the blood, in which their survival time is short (142).
Neutrophil Heterogeneity
In the past, polymorphonuclear neutrophils were thought to be a homogeneous population of end-stage cells incapable of protein synthesis and of essentially uniform size, granule content, and functional capability. Sabin first suggested potential heterogeneity among neutrophils when she reported that myelocytes were less motile than more mature neutrophils (141). A range of rates of motility among neutrophils from a single individual has been observed (143,144), and Harvath and Leonard suggest the existence of two neutrophil populations based on chemotaxis (145). Subsequently, several monoclonal antibodies were described that recognize subpopulations of neutrophils, including one that appears to recognize the classic NB1 (HNA-2a) neutrophil antigen and one that recognizes an activation epitope on CD11/CD18 (146,147,148). Another antibody, 31D8, appears to recognize a neutrophil subset that is more responsive to FMLP as determined by chemotaxis and respiratory burst activity (146). Neonates have a larger percentage of neutrophils that express low levels of 31D8 antigen (149). It has been reported that CR2 (CD21), the receptor for C3d, is present on immature neutrophils but not mature blood neutrophils (150,151). One report found that neutrophils from patients with localized juvenile periodontitis express CD21 (CR2) on their surface, whereas normal neutrophils do not (152).
Some studies of these different populations of polymorphonuclear neutrophils have been interpreted as reflecting maturation or environmental influences (153), in some cases possibly reflecting intravascular exposure to stimuli (154,155). The clinical significance of neutrophil subpopulations is unclear.
![]() Eosinophils
Eosinophils
Eosinophils (Fig. 14.1) exhibit the same maturation phenomena as neutrophils, with the exception that only one type of granule is recognized (83,91). In humans, these homogeneous granules appear to be formed throughout all the subsequent stages of maturation. Their contents are first seen as flocculent material in Golgi saccules, then in small vacuoles that condense to form the large homogeneous, dense granules. Subsequently, crystalloids develop, and the granules acquire an angular shape; the angular configuration predominates in the mature polynuclear forms (124). It is unusual to find more than two lobes in mature eosinophils, and the lobes are larger than those seen in neutrophils. The presence of eosinophils with more than two nuclear lobes suggests cell activation, as occurs in parasitic diseases. Eosinophilic granules are considerably larger than neutrophilic granules, appear somewhat refractile under the light microscope, and stain a bright yellowish red with Wright stain (102).
Human eosinophil granules contain eosinophil peroxidase (106), a heme protein that is distinct from neutrophil MPO (156,157); several cationic proteins, including eosinophil cationic protein (158,159); and eosinophil major basic protein (MBP). MBP constitutes a major proportion of total granule protein and damages several parasites such as schistosomes, Trichinella spiralis larvae, and trypanosomes as well as many types of mammalian cells, including respiratory epithelium (159). The granules also contain eosinophil-derived neurotoxin, the functions of which are less well understood (159). In some disease states, evidence for the involvement of eosinophils has been reported by the detection of MBP or erythropoietin at the site of pathology (160,161). Of the mature granulocytes, eosinophils display the most intense staining with peroxidase; the intensity of staining appears to be related to their basic protein content. Eosinophils are discussed more fully in Chapter 10.
![]() Basophils and Mast Cells
Basophils and Mast Cells
Basophils play an important role in allergic reactions. They express a unique surface antigen profile (162) and contain a variety of vasoactive and immunomodulatory chemicals that are released on activation. Basophils are distinguished by their large, coarse, purplish-black granules (Fig. 11.2A) that usually fill the cytoplasm and often obscure the nucleus. The granules are water-soluble and thus may be dissolved in the process of staining and washing; the cells may then appear vacuolated, with only a few or no basophilic granules remaining. On EM examination, a similar variation is noted in the appearance of the basophilic granules, possibly reflecting variable extraction of granule contents during preparative procedures (83,163,164).
The primary basophil granule is formed during the early cell stages (168). These primary granules are peroxidase-positive (105,107) and contain large amounts of heparin and histamine (107,135,166) (sulfated acid mucosubstance), features that are probably responsible for the affinity of the granules for basic dyes. These granules also contain the slow-reacting substance of anaphylaxis, kallikrein, an eosinophil chemotactic factor, and a platelet-activating factor (167). Charcot-Leyden crystals are also seen with both phase microscopy and EM; MBP is found in basophils as well as in eosinophils (87,165). A second, smaller granule that is bounded by a 5-nm membrane identical to that in the endoplasmic reticulum, Golgi apparatus, and mitochondria has also been described (166). These granules are peroxidase-negative (105).
Similar but somewhat larger cells found in the tissues are mast cells. Mast cells normally do not circulate in the blood (168). Although these cells resemble basophils in their metachromasia, acid nature, and content of histamine and heparin, they contain hydrolytic enzymes, 5-hydroxytryptamine (166), and serotonin (107,169), which basophils do not. Also, the ultrastructure of their granules is different in humans and guinea pigs (169). Basophils are discussed more fully in Chapter 11.
![]() Macropolycytes
Macropolycytes
Macropolycyte is the name applied to giant polymorphonuclear neutrophils with a diameter greater than 16 μm and with 6 to 14 nuclear lobes (170). Such cells are seen only occasionally in healthy subjects (1.3%), but they are found in approximately 5% of people with infections of various types or with intoxications, usually in association with a neutrophilic leukocytosis and myelocytes in the blood (170). Macropolycytes are commonly seen in association with folic acid or vitamin B12 deficiency, as well as in patients recovering from the pancytopenia that attends treatment with cytotoxic agents, especially hydroxyurea.
|
|
|
Figure 9.10. Granulocytes and sex chromatin patterns. Two cells on the left show the characteristic drumsticks found in the female subjects. The thin strand of chromatin joining the head to a nuclear lobe can be seen clearly. In the two cells on the right, small clubs, such as may be seen in male subjects, should not be confused with drumsticks (Wright stain, ×1,300). |
Some authors describe cells with hypersegmented nuclei but of a normal size and call them polycytes (128) or polylobocytes (131); similar cells with complex nuclei but without hypersegmentation are called propolycytes (128). The latter forms are seen in approximately 10% of patients recovering from leukocytosis with a marked shift to the left and appear in increasing numbers when anticoagulated blood is allowed to stand in vitro (128,171).
The mechanism of macropolycyte formation is unknown, but one suggestion is that the skipping of one of the usual cell divisions that occurs during maturation results in a hypersegmented cell (172).
![]() Genetic Sex as Indicated by Leukocytes
Genetic Sex as Indicated by Leukocytes
Only one X chromosome is essential to the normal activity of a cell; the other in the normal XX female remains unextended and thus is visible as a chromatin body. Sex chromatin (Barr) bodies are present in 80 to 90% of the somatic cells of the normal female subject. The sex chromatin body of the neutrophil of females is a small mass, usually adjacent to the nuclear membrane, that stains deeply with hematoxylin, Feulgen reagent, and thionine and is approximately 0.7 to 1.2 μm in diameter. It takes the form of a drumstick projecting from one of the nuclear lobes of approximately 2 to 3% (extreme range, 1 to 17%) of the segmented neutrophils in the blood (173). They are well-defined, solid, round projections of chromatin connected to a lobe by a single, fine chromatin strand (Fig. 9.10). They must be distinguished from small clubbed or racket-structured, nonspecific nodules that may be smaller or larger as well as irregular in shape or lacking in chromatin, as well as from small (minor) lobes attached to the rest of the nucleus by two strands. Confirmation of the X chromosome in the drumstick has been provided by in situ hybridization (174). Sessile nodules are equally gender-specific but are more difficult to recognize (163). Drumsticks are not found in normal male subjects (175).
The number of chromatin bodies seen in a cell is one less than the number of X chromosomes present. With the increased numbers of X chromosomes found in certain disorders of human development, the number of Barr bodies and drumsticks increases, and isochromosomes formed by duplication of the long arms of the X chromosome give rise to larger drumsticks than are found in the normal female subject (176). Drumsticks or sessile nodules are seen in chromatin-positive male patients with Klinefelter syndrome and are absent in chromatin-negative female patients with Turner syndrome. Eosinophils and probably basophils also have drumsticks. Drumsticks may be difficult to find in the presence of a marked shift to the left. Drumstick counts are reportedly low in the leukocytes of patients with chronic myelocytic leukemia, paralleling the low leukocyte alkaline phosphatase and catalase concentrations (177). Double drumsticks (175) or a sessile nodule plus a drumstick in the same neutrophil are rare (178).
![]() Differentiation of Various Types of Cells
Differentiation of Various Types of Cells
Monocytes are described in detail in Chapter 12; lymphocytes and plasma cells are described in Section 4. The morphologic characteristics of the various leukocytes are summarized in Tables 9.3 and 9.4 to facilitate comparison. Nucleated forms of the erythrocyte series are easily distinguished from most of the leukocytes by their lack of cytoplasmic granules. In Romanowsky-stained films, confusion arises only in the immature (blast) stage. Even then, the more granular and clumped nuclear chromatin may help to identify the pronormoblasts. Polychromatophilic normoblasts at times may be confused with plasma cells or lymphocytes. In the polychromatophilic normoblast, however, the nucleus is more centrally placed than that of the plasma cell or lymphocyte, the cytoplasm is blue-pink, and the cell border may be irregular. The mature lymphocyte is characterized by coarse, clumped chromatin in a nucleus that is eccentrically placed in sparse cytoplasm; a few granules also may be present. Deep blue cytoplasm in a cell with an eccentric nucleus containing coarse, clumped chromatin is the hallmark of the plasma cell.
The nucleated cells of the red cell series are nonmotile; in wet films, they have a rounded, distinct border with homogeneous, nongranular yellowish cytoplasm. The nucleus is round or oval and is centrally placed. The chromatin arrangement gives the nucleus a vesicular appearance; in early forms, one or two large nucleoli are present. In supravitally stained preparations, no neutral red bodies are seen, but many coarse, rod-shaped, and coccoid mitochondria are scattered diffusely in the cytoplasm.
Students will find it valuable, when learning to identify the various cells of the blood and bone marrow, to seek each of the morphologic criteria listed in Tables 9.3 and 9.4 in systematic fashion. By doing so, they will acquire the habit of seeing all that they are viewing and, in time, learn to identify cells because of a number of characteristics perceived unconsciously. The actual identification of cells regarding which some doubt remains can be made only by weighing the evidence for and against each type being considered. One fact to keep in mind is that practically no characteristic of a cell is entirely specific. Thus, a perinuclear clear zone is sometimes seen in cells other than plasma cells, and a rosette of neutral red bodies has been observed in many types of blood and connective tissue cells other than monocytes.
![]() Differential Cell Counting and Normal Values for Leukocytes
Differential Cell Counting and Normal Values for Leukocytes
Differential cell counting is the enumeration and classification of the leukocytes seen on the blood smear. The usual procedure is to count at least 100 consecutive leukocytes in an area of good cell distribution. A uniformly thin smear of blood on a cover glass is the best preparation for such examination.
Distributional errors are reduced as more cells are counted. Confidence tables or curves can be used to estimate the probable error of a differential count when various numbers of cells are counted. Table 9.5 shows 95% confidence limits. Clearly, as more cells of a given type are counted and as the total number of cells enumerated increases, the accuracy of the differential count is greater. Thus, if 200 cells are counted, and a frequency of 70% is found for a given cell type, the true value can be expected to lie between 63.5 and 76.5% for 95% of such counts. If a subsequent 200-cell differential count gives a figure of 80% for that cell type, the difference is probably real, whereas if only 100 cells have been counted, the difference would probably not be significant. Even so, when dealing with cells present only in small numbers (such as eosinophils or basophils in the usual smear), the values obtained from the differential count provide only a gross estimate of cell frequency. For more accurate enumeration of these cell types, absolute counting methods have been developed.
From the total leukocyte count and the differential count, the absolute concentration of each leukocyte type can be calculated. The accuracy of the result depends on the validity of the total leukocyte count and the differential count. With automatic cell counters, the major component of error now lies in the differential count.
Normal values for absolute leukocyte concentrations obtained by using a Coulter counter and differential counts are shown in Table 9.6. Similar values have been reported with the use of other methods (179). Some variation is evident in values obtained in different population groups and appears to depend on age, sex, pregnancy, time of day, activity level, and other factors (180). Racial variation has been reported, especially in Ethiopian Jews (181) and black Africans, who have significantly lower neutrophil and monocyte counts (182). Lower counts are less evident in black Americans (129) and in Africans eating a Western diet (180).
The absolute leukocyte concentration provides a more accurate picture than the differential count because each leukocyte type is a separate cell system with its own functions, control mechanisms, and responses to disease processes; for example, a patient with chronic lymphocytic leukemia whose total leukocyte count is 100 × 109 cells/L, 7% of which are neutrophils and 93% are lymphocytes, does not have granulocytopenia. With a blood neutrophil concentration of 7.0 × 109 cells/L, the problem is lymphocytosis.
Various systems for differential counting have been used (Fig. 9.11) (183). Arneth, for example, painstakingly recorded and tabulated from left to right the number of neutrophilic leukocytes with 1, 2, 3, etc., lobes and made other subdivisions (138). The term shift to the left is derived from this practice and indicates an increase in the proportion of cells with only one or few lobes, whereas shift to the right represents an increase in the proportion of multisegmented forms.
From a clinical viewpoint, it is useful to determine whether young forms of neutrophils (band forms and younger) are increased and whether the proportion of multinucleated forms is increased. An increase of younger forms (band cells, metamyelocytes, and myelocytes) (shift to the left) suggests increased release of young neutrophils from the bone marrow, which is seen in association with acute infections (134) and inflammation. If a shift to the right is suspected, a neutrophil lobe count may be useful. This process involves counting the total number of nuclear lobes in 100 or 200 neutrophils, calculating the average lobe number/neutrophil, and comparing the results with normal values. The chief difficulty associated with this count is clear definition of what constitutes a separate lobe (see “Band Neutrophils” earlier in this chapter). If complete separation of nuclear lobes with or without a connecting filament is the definition used, the normal mean neutrophil lobe count is 2.04, with 95% of normal values falling between 1.66 and 2.42. An increase in mean neutrophil lobe count suggests vitamin B12 or folic acid deficiency, congenital hypersegmentation of neutrophils, or renal disease (133). A ratio of five-lobed to four-lobed polymorphonuclear cells that is greater than 0.17 is said to be associated more regularly with B12 deficiency than is an increase in mean nuclear lobe count (131).
Alterations in the total number of leukocytes and in their relative proportions are significant as measures of the reactions of the body to noxious agents. The reactions of leukocytes in association with certain diseases are discussed later in this book, as is the presence of abnormal inclusions, such as toxic granulation, Döhle bodies, and various inherited abnormalities in leukocyte morphology.
|
Table 9.4 Morphologic Characteristics of Leukocytes (Supravital Stain) |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
An additional type of differential cell count in common use is the histochemical, semiquantitative estimate of the leukocyte alkaline phosphatase content of neutrophils (see Chapter 1) (117).
![]() Neutrophil Kinetics
Neutrophil Kinetics
The importance of leukocytes in the defense of the organism is well known. Basic to their roles are cell multiplication, maturation, storage, and delivery to the tissues and sites of infection or cell damage. These processes are called leukocyte kinetics and are different for each leukocyte type. To simplify the discussion, each type of leukocyte is considered as a separate system, but these systems constantly interact and complement one another in the defense of the body.
Neutrophil Kinetics in the Adult
In Chapter 5, the process of mitotic cell division, the cell generation cycle, and the origin of neutrophils and other cell types from a multipotent hematopoietic stem cell were discussed. The multipotent hematopoietic stem cell is thought to produce a committed stem cell from which the myeloblast and monoblast are formed (86); from them, the neutrophilic and monocytic series are derived. The production, kinetics, and lifespan of the neutrophil have been the subject of a number of reviews (184,185,186,187,188,189,190,191,192,193,194,195,196,197,198). A model of these processes in adult humans is shown in Figure 9.12. The neutrophil system appears to be incompletely developed in premature babies and in early neonatal life; this topic is discussed in the section, “Neutrophil Kinetics in the Fetus and Newborn” later in the chapter.
Mitotic and Maturation Compartments
Neutrophil production in normal adult humans appears to take place only in the bone marrow. The life cycle of the neutrophil can be divided conveniently into bone marrow, blood, and tissue phases. The assumption is that cells move through the system in an orderly manner as if in a pipeline; this view is supported by the progressive movement of isotopic tracers (115,195,200,201) and azurophilic granules (83,93,101) through the system.
The myeloblast, promyelocyte, and myelocyte are capable of cell division, as judged by direct observation in cultures (91) and by their ability to incorporate 3H-TdR into their nuclear DNA (201). These forms, therefore, constitute the mitotic compartment (Fig. 9.12). Simultaneously, they undergo differentiation, as evidenced by the appearance of azurophilic and specific granules in their cytoplasm. The more mature forms of the neutrophil series (metamyelocyte, band, and polymorphonuclear neutrophil) are usually considered incapable of cell division (except perhaps in unusual circumstances) (202) and do not incorporate 3H-TdR into their nuclei, but they do exhibit continuing maturational changes and thus constitute the maturation compartment. From the maturation compartment, cells flow into the blood and are distributed in two sites: the circulating blood granulocyte pool (CGP) and the marginal granulocyte pool (MGP). Cells in these two pools are in constant equilibrium. Eventually, the cells move through vessel walls to enter the tissues. The exact nature of the MGP is not clear. In the past, it was felt to represent transient adhesion to and rolling along the surface of endothelial cells, primarily in postcapillary venules. However, subsequent studies of a patient with leukocyte adhesion deficiency-2 (LAD-2) demonstrated the presence of a marginating pool (203), although the MGP in this patient with LAD-2 was reduced. Patients with LAD-2 lack the ligands for the selectins CD62P and CD62E and have a marked decrease in neutrophil rolling in postcapillary venules. Normally, the MGP is approximately equal in size to the CGP. Studies of this patient suggested that approximately 20% of neutrophils are in a selectin-independent MGP and approximately 30% are in a selectin-dependent MGP (203). Surprisingly, this patient’s neutrophils had a shorter than normal half-life in the circulation with an increased turnover rate. The inability of CD18 or CD62L antibodies to shift neutrophils from the MGP to the CGP also suggests that the transient adhesion to endothelial cells manifest as rolling does not account for the MGP (204,205).
In this model, cell production can be estimated either by assessing the production rate in the mitotic compartment or by measuring cell flow through subsequent stages, such as the blood. Accuracy of these measurements is facilitated if the system is studied in the steady state when compartment sizes are constant and cell flow reflects net production and destruction (191).
|
Table 9.5 95% Confidence Limits for Differential Leukocyte Counts |
|||||||||||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||||||||||
If one assumes a steady state in a scheme such as that shown in Figure 9.7, the flow of cells out of any pool (Kout) is equal to the flow of cells into that pool (Kin) plus any cells produced in the pool (Kb); thus,
Kout = Kin + Kb
Clearly, in pools other than those in the mitotic compartment, Kb is 0, and measurements of cell flow (Kin or Kout) equal effective cell production, provided no cell death occurs within the pool.
Production in the Mitotic Compartment
Because myeloblasts, promyelocytes, and myelocytes constitute approximately 0.9, 3.3, and 12.7% of the marrow cells, respectively, it has been assumed that the system has four or five divisions (197). From a review of blood neutrophil radioactivity curves obtained after di-isopropylfluorophosphate (DF32P) injection into humans, investigators raised the possibility of at least three divisions at the myelocyte stage (197). Another suggestion generated from marrow differential counts was that only four or five divisions occur in the entire neutrophil proliferation scheme (197). This supposition agreed with results from experiments in dogs (206) and with data from model studies (207). In contrast, studies of myeloid islands in the rat thymus provided evidence for seven divisions during granulocytopoiesis: one in the myeloblast stage, two in the promyelocyte stage, three in the myelocyte stage, and a final one in the metamyelocyte stage (136,208).
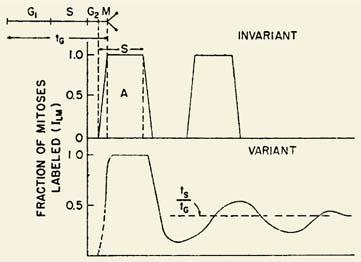
Calculations made from mitotic index (MI) data (189,209,210,211) provide estimates of cell generation time (tg) and pool turnover time. MI (209,210,213) is defined as
|
Table 9.6 Normal Blood Leukocyte Concentrations (95% Confidence Limits) |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Figure 9.11. Several classifications of neutrophils. Note that all the classifications agree on a common dividing line between mature and immature cells. The Schilling classification further subdivides only the immature cells. The Cooke and Ponder and the Arneth classifications further subdivide only the more mature cells. (From Haden RL. Qualitative changes in neutrophilic leukocytes. Am J Clin Pathol 1935;5:354, with permission.) |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
MI = Nm/N
in which MI is the mitotic index for any morphologic cell pool, Nm is the number of mitoses in that pool, and N is the total number of cells in the pool. MI can also be expressed as the ratio of the time spent in mitosis (tm) to the cell generation time (tg):
MI = tm/tg
By combining both definitions,
MI = Nm/N = tm/tg
By providing determined values for MI and mitotic time (tm) in the last equation, the generation time (tg) for a particular cell pool can be approximated. From tg and the pool size (N), the birth rate, Kb, can be obtained if all cells in the pool are in cycle, because each mitosis gives rise to one new cell:
Kb = N/tg
In effect, the cell birth rate is equal to the number of mitoses occurring per unit time (t), or
Kb = Nm/t
Although the concept is simple, several problems arise (211,214). A major problem is that the morphologic boundaries of most cell pools are not clearly delineated in terms of the cell cycle (214). For example, to calculate cell production in the myelocyte
P.188
pool, it must be assumed that all myelocytes are destined to divide; that is, no cells are recognized as myelocytes that are not going to divide again. Because the daughter cells of the last myelocyte mitosis almost certainly do not suddenly become metamyelocytes on completion of division, N in the preceding equation will be erroneously large, and thus estimates of tg will be erroneously long.
|
|
|
Figure 9.12. Model of the production and kinetics of neutrophils in humans. The marrow (557) and blood compartments (148) are drawn to show their relative sizes. In the lower one third of the figure, the compartment transit times as derived from diisopropylfluorophosphate (DF32P) studies (139,148) and from tritiated thymidine (3H-TdR) studies (152,557) are compared. CGP, circulating granulocyte pool; MB, myeloblast; MGP, marginal granulocyte pool; myelo, myelocyte; pro, promyelocyte. |
If the fraction of nonmitotic cells in the myelocyte population were known, the calculations could be corrected for this error, as has been attempted (187). A second major problem is the fact that values for the MI have varied considerably (209,211,212,215,216,217,218). In addition, a considerable diurnal variation exists in the MI in humans, as well as in animals (211,218,219,220).
Finally, to calculate the absolute neutrophil production rate (in cells per unit of time), the size of the marrow mitotic compartment must be known. Methods for measuring the sizes of marrow myeloid pools have been developed (221,222,223,224) (see “Size of Marrow Compartments and Their Morphologic Subdivisions,” later in the chapter), but to date no one has measured these sizes and MI in the same animal at the same time and then calculated neutrophil production rate. Nevertheless, values for the MI for each of the neutrophil precursors capable of mitosis have been determined (211,212), and within the assumptions inherent in such calculations (214), neutrophil production has been estimated (193,210).
Similar calculations of neutrophil production can be made from 3H-TdR–labeling index data. After flash labeling with 3H-TdR, autoradiographs of the bone marrow are obtained, and the proportion of nucleated cells that have incorporated the label into their nuclei is determined (188,194). This labeling index (LI) represents the ratio of labeled cells, *N, or cells in DNA synthesis (Ns) to total cells (N) of a defined morphologic type:
LI = *N/N = Ns/N
The LI can also be defined in terms of DNA synthesis time (ts) and the cell generation time (tg), because 3H-TdR is taken into the cell only during the period of DNA synthesis; thus,
LI = ts/tg
By combining both definitions,
LI = Ns/N = ts/tg
and from determined values for LI and tg, the generation time and turnover of a given cell population can be estimated. As with MI data, birth rate is a function of the population turnover time, which can be approximated from the generation time or time spent in various phases of the cell cycle:
Kb = N/tg = Ns/ts
Some of the same problems arise with the 3H-TdR LI that are encountered in the use of the MI (214,225). In addition, the use of labeled compounds raises questions of label reuse (226,227) or elution (225) and perturbation of the cell population by the compound (185,206,228,229,230) or by its radioactivity (225,231,232).
The LI reported for humans is myeloblast, 0.85; promyelocyte, 0.65; and myelocyte, 0.33 (188). Somewhat different values have been reported in dogs (194) and rats (212). By using the LI for humans and a value for ts of 5 hours (based on studies in dogs) and by determining relative compartment sizes for each cell type from the bone marrow differential count, the relative birth rates (Kb) of cells have been calculated (187,212).
Some authors have found good agreement between neutrophil production as calculated from the MI and the LI (212), but considerable discrepancy has been reported by other authors (187). This difference may result from the fact that the MI values obtained were low, the studies were done in different subjects at different times, and too small a value for ts was used in the calculations.
The turnover time of a labeled compartment and neutrophil production rate also may be estimated by measuring the grain count halving time (199). The generation time is derived only if each cell in a given class divides and if no label feeds into the compartment from a labeled precursor class or as a result of label reuse (233). If any of these criteria are not met, the half-time for grain count decrease is longer than the true value, and the estimate of generation time is only a maximal value. Additional disadvantages of this method are that at least several bone marrow samples distributed throughout several half-times are needed, and that grain counting is extraordinarily tedious and subject to considerable error. Nevertheless, estimates of compartment turnover time have been made with this method by using 3H-TdR (199) and radiosulfate (124).
After flash labeling with 3H-TdR, the cohort of cells labeled during DNA synthesis may be followed as it subsequently enters mitosis, and the time course of labeled mitoses can be recorded (233,234). Theoretically, such curves should permit measurement of the post-DNA synthesis gap (G2), mitotic time (tm), DNA synthesis time (ts), cell generation time (tg), and pre-DNA synthesis gap (G1) (see Chapter 5 and Fig. 9.13). In actual practice, biologic variation rounds off the percentage of labeled mitosis curves, and rapid damping of the waves of cells passing through mitosis (Fig. 9.13) renders such measurements less precise than ideal. However, estimates of myeloid DNA synthesis time obtained with this method are approximately 11 to 13 hours in humans (233,234) and are in good agreement with estimates made in gastrointestinal mucosal cells. From the level of the damped plateau reached after a few hours, the ratio of ts to tg can be obtained (Fig. 9.13), and the generation time can then be calculated. If the generation time and compartment transit time are presumed to be the same or if the proportion of cells in a compartment that is actively proliferating is known, the neutrophil production rate can be calculated.
Neutrophil Production as Measured by Cell Flow in Other Compartments
Another method for approximating neutrophil production involves following the appearance of 3H-TdR–labeled cells in the metamyelocyte compartment. Because metamyelocytes do not divide or take up 3H-TdR, the appearance of labeled cells in this compartment should reflect the flow of cells into it from the myelocyte compartment; in the steady state, this influx of cells should also reflect the turnover of the metamyelocyte compartment and thus cell production. Approximately 3 hours pass after the injection of 3H-TdR before label appears in metamyelocytes both in dogs (194) and in humans (233); this time interval is the minimum time for myelocytes taking up the label to pass through G2 and mitosis and become metamyelocytes. After this lag, the rate of labeled cell inflow into the metamyelocyte compartment is approximately 3 to 5%/hour in both species. In the dog, cell inflow into the metamyelocyte compartment measured in this fashion is less than 50% of that calculated from LI data (194), suggesting the existence of a major component of ineffective granulocytopoiesis, a myelocyte sink, in the normal animal. However, similar calculations in humans do not confirm the findings of such studies (187). The resolution of this enigma requires the simultaneous measurement of cell production by using several methods in the same animal at the same time.
|
|
|
Figure 9.13. The percentage of labeled mitoses in the course of cell generation. At top of the diagram is the theoretical configuration that would be seen if cells flowed through the proliferation cycle with no variation. The effects of moderate variation in time spent in the several cycle phases on the percent labeled mitosis curve are shown at the bottom. G1 is the pre-DNA synthesis resting phase (gap), S is the DNA synthetic period, G2 is the post-DNA synthesis gap, and M is mitosis. tS is the time spent in S, and tG is the duration of the entire generative cycle. (From Cronkite EP, the Brookhaven National Laboratory, and the National Cancer Institute. Kinetics of granulocytopoiesis. Natl Cancer Inst Monogr 30, Human Tumor Cell Kinetics 1969;51.) |
In a similar manner, the marrow production of neutrophils has been estimated from the size of the postmitotic maturation storage compartment and the compartment turnover time (212,222,235). The compartment size is calculated from the marrow neutrophil-to-erythroid (NE) ratio (determined in marrow sections) and the mean normal marrow normoblast pool size (calculated by multiplying the ratio of subjects’ erythroid iron turnover value over the mean normal value for this determination by the mean normal erythroblast population) (222). The compartment transit time is estimated by injecting 3H-TdR and noting the time required for labeled neutrophils to appear in the blood. By this method, marrow neutrophil production in humans has been calculated to be 0.85 × 109cells/kg/day in the normal steady state.
Neutrophil production also can be approximated by measuring the flow of cells through the blood, the blood granulocyte turnover rate (GTR). DF32P binds irreversibly with a number of esterase enzymes and has been shown to label neutrophils primarily (236,237). By means of this agent, a subject’s own cells can be labeled, and the total blood granulocyte pool (TBGP) and the rate of disappearance of labeled neutrophils from the blood can be determined (192). Similar measurements can be made by transfusing 3H-TdR–labeled cells from a suitable donor (222). Because neutrophils leave the blood in a random manner (exponential disappearance curve), the GTR is calculated as follows from the TBGP and the t1/2:
GTR = 0.693/t1/2 TBGP
in which 0.693 is the natural logarithm of 2 and t1/2 is the blood neutrophil half-disappearance time.
An important point to keep in mind is that all of these methods assume that the system is in a steady state during the entire course of the measurements. If neutrophil death in the marrow is not significant, the blood GTR equals total neutrophil production. If neutrophil death in the bone marrow is significant, the blood GTR measures effective neutrophil production, and the difference between this determination and total neutrophil production is ineffective granulocytopoiesis. Measurements of neutrophil production by compartment turnover methods have given values ranging from 62 to 400 × 107 neutrophils/kg/day in humans (Table 9.7) (222,238) and 150 to 560 × 107 neutrophils/kg/day in dogs (222,235,239).
Of the methods for assessing neutrophil production just described, only the measurement of blood neutrophil turnover rate with DF32P or 3H-TdR can be performed easily enough to be of use in studying groups of patients in a clinical setting, and even this is possible in only a few research centers.
Size of Marrow Compartments and Their Morphologic Subdivisions
In all of the procedures described, with the exception of DF32P and 3H-TdR blood kinetic measurements, the number of marrow myeloid cells under study must be known to calculate neutrophil production. In the absence of such data, only calculations of relative cell production are possible (187). Direct measurements of the volume and cellularity of the marrow (isolation of skeleton and cell counting or bone biopsy and radioactivity measurement) have been made in dogs (222), rats (240), mice (241), and guinea pigs (242,243).
|
Table 9.7 Blood Neutrophil Kinetic Parameters in Normal Humans |
|||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||
Several methods for estimating marrow myeloid mass have been devised, but probably the best data are those derived by using marrow NE ratios and measurement of marrow erythroblast mass by the iron dilution technique as originally suggested by Suit (244). In early studies by Donohue et al. (23,224), marrow erythroblast radioactivity and NE ratios were counted in marrow suspension smears. Corrections were made for incomplete iron localization in the erythron and for an estimated 30% cell destruction that occurred in preparing the marrow suspensions. In later studies by Harker (245), the NE ratios and radioactivity measurements were made on marrow sections in the hope of avoiding the problem of possible cell destruction. Values for marrow erythroid mass were somewhat lower than those reported by Donohue et al. (223,224), but they agreed with results obtained when no correction for cell destruction was made. By using marrow sections and the 59Fe isotope dilution technique, investigators have directly determined the marrow myeloid mass in the dog (222). In humans, however, direct measurement of the marrow normoblast mass has not been feasible in normal subjects, so marrow normoblast mass was estimated from erythron iron turnover values (222,246). The mean values obtained in 13 normal subjects are presented in Figure 9.12.
Transit Time through the Nondividing Maturation Pool
After injection of a pulse label of 3H-TdR or radiophosphate, a delay of several days occurs before labeled segmented neutrophils appear in the blood. This interval represents the minimum time from DNA synthesis in the last myelocyte generative cycle until the cell has matured into a segmented neutrophil (or band form) and is released into the blood. In patients in a normal steady state, this minimum transit time or emergence time was between 96 and 144 hours (247,248); in patients with infection, it was as short as 48 hours. Emergence time in dogs is 2 to 3 days (249); in the rat, it is 36 to 42 hours (139).
The mean value for myelocyte-to-blood transit time after DNA labeling (defined as the time from 3H-TdR or radiophosphate injection to the peak of the blood granulocyte radioactivity curve) was 6 to 9 days in hematologically normal convalescent patients and 7.2 days in eight normal subjects (221). In contrast, studies in which DF32P was injected intravenously into normal volunteers (prisoners) led to the conclusion that the mean promyelocyte-to-blood transit time was 11.4 days, with a myelocyte compartment transit time of 2.9 days (186,197)—in other words, an 8.5-day myelocyte-to-blood transit time. Results of studies involving the simultaneous use of 3H-TdR and DF32P in the same subjects suggest that the discrepancies reflect differences between normal ambulatory subjects and hematologically normal convalescent patients (250). In dogs, the average myelocyte-to-blood transit time was 5 days, as measured with both 3H-TdR and DF32P (250).
Neutrophil Release from Marrow into Blood
After 3H-TdR injection and at the time of first seeing label in band and segmented neutrophils in the bone marrow, some labeled cells are also seen in the blood (187). Authors have suggested that the release of band or segmented neutrophils from the marrow does not follow a strict pipeline or first-in, first-out sequence (187,247). However, it is not clear whether these observations reflected variance around a mean transit time (197,249,251) or random release of mature neutrophils from the marrow (247). Findings in dogs strongly favor the mean transit time concept rather than random release (249). In fact, because the ratio of band to segmented cells is much higher in marrow than in blood, selective release of segmented cells must occur.
The mechanisms controlling the release of marrow cells into the blood are only partially understood. Endotoxin disturbs the relationship between marrow sinus endothelial cells and the adventitial (reticular) cells (252,253), which usually cover approximately 60% of the extraluminal sinusoid surface (254). Endotoxin induces a reticular cell movement away from the endothelium, thus facilitating hematopoietic cell contact with and migration between the endothelial cells (252,254). Findings of in vitro studies of factors influencing marrow granulocyte migration through membranes demonstrate that pore diameter, morphologic age of cells, and the presence of a chemical attractant may be important in marrow cell release (255). Thus, immature granulocytes (myeloblasts and promyelocytes) could not penetrate membranes with pores smaller than 8 μm and were not responsive to chemoattractants. Mature granulocytes (band and segmented) could penetrate membranes with pores as small as 1 μm, and egress was accelerated by increasing pore size and by use of a chemoattractant. Myelocytes and metamyelocytes exhibited intermediate activity. Integrins and immunoglobulin (Ig) superfamily members appear to play an important role in the organization of the bone marrow microenvironment, and stem cell factor alters the avidity of α4β1 and α5β1 integrins on hematopoietic cell lines for their ligands (256). A number of mediators of neutrophil release from the bone marrow have been identified, including tumor necrosis factor (TNF)-α, TNF-β, granulocyte colony-stimulating factor (G-CSF), granulocyte-macrophage colony-stimulating factor (GM-CSF), IL-8, and C5a (203,257,258).
Kinetics in the Blood
Di-isopropylfluorophosphate Studies
With the development of the DF32P labeling technique, in which neutrophils are labeled in vitro with DF32P and returned to the donor, two pools of neutrophils in the blood have been demonstrated: a CGP and an MGP (192,259,260). In normal humans, neutrophils in these two pools are in constant equilibrium, and the pools are of approximately equal size (Table 9.7). The CGP is calculated from the blood leukocyte count and the blood volume. The TBGP is measured by the dilution of DF32P-labeled neutrophils after their reinjection (259). MGP is the difference between the TBGP and the CGP.
Brief exercise or epinephrine injection increases the size of the CGP by approximately 50%, but the TBGP is unchanged; the neutrophilia produced reflects a demargination of cells that lasts only a few minutes (259). The location of at least some of the marginated cells is thought to be along the walls of small blood vessels, primarily postcapillary venules, in many body tissues, where neutrophils roll along the vessel wall in loose contact with the endothelial cell surface (261). The distribution of cells in the CGP and MGP can be altered by other means. For example, transient neutropenia was noted 90 minutes after endotoxin injection, but the TBGP was not significantly increased; thus, essentially a shift from CGP to MGP occurred. At the end of 5 hours, the TBGP increased significantly as a result of an outpouring of cells from the bone marrow, and the CGP and MGP were of approximately equal size (260). The administration of steroids also produced an increase in the size of the TBGP, in part for the same reason but also because of decreased outflow into the tissues (262). A rapid transient increase in the marginating pool has been observed during hemodialysis when cuprophane membranes are used (263,264). Activation of the complement system by contact with the dialyzer membrane was found to produce inflammatory mediators (C5a being a major contributor) that stimulated an increase in neutrophil margination manifest both by an increase in rolling along small vessel endothelium and by the formation of homotypic aggregates of neutrophils with sludging in small vessels (265,266,267,268,269). This neutrophil margination was transient, being maximal by approximately 15 minutes and resolving by 1 hour of dialysis.
After the return of DF32P-labeled neutrophils to their donor, the disappearance of labeled cells from the blood follows a single exponential curve with a half-disappearance time (t2) of approximately 7 hours in most normal subjects (186,192,221,270,271,272,273). This finding implies that neutrophils are destroyed or leave the blood randomly rather than according to their age (senescence), as is the case for erythrocytes and platelets (192). Support for this theory is found in the fact that 3H-TdR–labeled neutrophils appear in the blood and in saliva almost simultaneously (142).
Studies involving leukapheresis (274) and other experiments (275) reveal that neutrophils that have crossed the endothelial barrier between blood and tissues probably do not re-enter the circulation, at least not in significant numbers. The number of neutrophils that pass through the blood each day has been estimated at 62 to 400 × 107 cells/kg/day (Table 9.7)—that is, the GTR. In 1964, it was proposed that neutrophils became senescent and developed pyknotic nuclear lobes over time, this process truncating the exponential curve of disappearance of DF32P-labeled neutrophils (142). Although this concept seemed plausible, no truncation of DF32P curves has been noted. Similar blood kinetic results have been obtained with the use of 3H-TdR and autoradiographic techniques (276). More recent studies describe the occurrence of apoptosis in neutrophils, and this may be an important mechanism of senescence in some circumstances.
51Cr Studies
Leukocytes can also be labeled in vitro with radiochromate (51CrO4) and then returned to the body (277,278,279). Unlike DF32P, which labels primarily granulocytes (235,237,276), leukocyte types other than neutrophils are heavily labeled by the 51Cr technique (248,277,279,280,281,282). Under some circumstances (such as chronic lymphocytic leukemia), this capacity is advantageous because the distribution and turnover of leukocytes other than neutrophils can be followed (277,280,282,283). In addition, 51Cr is a γ-emitting isotope, so external counting techniques may detect sites of sequestration or destruction.
Results of studies of the infusion of autologous, radiochromate-labeled leukocytes into hematologically normal subjects are more variable than those using the DF32P method. The proportion of cells recovered in the blood is sometimes less than that associated with DF32P-labeled neutrophils (279), and complex curves are often noted (283). Furthermore, there is no general agreement on t1/2values; most investigators report values similar to those obtained with DF32P (248,277,279,280,284), but values two to three times longer have been described (285,286). Several factors may explain these differences, but a major one is probably 51Cr labels of multiple cell populations. No significant organ sequestration of labeled leukocytes has been detected in normal subjects (279).
111In Studies
After the evaluation of a variety of other radioactive agents for labeling leukocytes in vitro (287), 111In chelated with 8-hydroxyquinoline (oxine) was thought to be promising for evaluating the in vivo localization of abscesses. The 111In-oxine complex labels blood cells effectively (95% uptake in 15 minutes) and exhibits minimal elution (<1% released in 2 hours) (288). This label is nonspecific, however, in that it binds to all cell types; thus, cell separation and purification are necessary before labeling occurs (289). Nevertheless, the labeling of autologous neutrophils (or leukocyte suspensions containing mostly neutrophils) and their reinjection is sometimes useful in localizing abscesses in humans, although much of the radioactivity initially localizes in the lungs and subsequently appears in the spleen and liver (288).
Apoptosis
Neutrophils undergo apoptosis in tissue culture (approximately 50% of cells in 24 hours); this process is accompanied by a fall in intracellular pH (290,291,292). Apoptosis is manifest by characteristic morphologic changes with nuclear condensation and the formation of pyknotic nuclei. Importantly, the cell remains intact and does not release its potentially toxic contents before it is phagocytosed by macrophages and removed. The addition of G-CSF, GM-CSF, or IL-1 delays but does not prevent this process (290,293). Macrophages phagocytose senescent (apoptotic) neutrophils (as observed more than a century ago), and this is probably one fate of the short-lived blood neutrophil (291,292,294,295,296,297). Interestingly, glucocorticoids inhibit neutrophil apoptosis (298), but they induce it in susceptible lymphocytes. Fas (CD95) is expressed on neutrophils and is capable of inducing apoptosis as it does in other susceptible cells (298,299). Neutrophil apoptosis is reversibly inhibited by hypoxia, possibly playing a role in neutrophil survival at sites of inflammation (300). Neutrophil apoptosis is also delayed in pregnancy, and both estradiol and progesterone can delay neutrophil apoptosis in men and women (301,302). Neutrophils may also undergo cell death by another mechanism that is caspase-independent, is not associated with DNA laddering, and appears to depend on mitochondria-derived reactive oxygen metabolites (303).
Migration into Tissues and Sites of Destruction
At a local site of tissue damage or infection, adherence of neutrophils to the endothelial cells of the vessel wall and their subsequent migration into the tissues can be seen within minutes. After initial adherence, neutrophils project microscopically visible pseudopods between the endothelial cells and force a passage between them (304,305). This directed movement, chemotaxis, is induced by the binding of a variety of chemoattractant molecules, such as n-formyl peptides (e.g., FMLP), a cleavage product of the fifth component of complement (C5a), leukotriene B4, and platelet-activating factor, to specific membrane receptors. Further migration is then delayed by the basement membrane and periendothelial cells, and the neutrophils may move parallel to, but beneath, the endothelium until a passage into the surrounding connective tissue is found. Once neutrophils leave the blood, they do not return in significant numbers (275,306).
The sites into which neutrophils normally disappear are poorly understood. Labeled blood neutrophils are found in saliva (139), but loss into saliva may reflect subclinical infections, because few if any cells are found in the salivary ducts (307); the rate at which granulocytes enter the oral cavity has been correlated with the degree of gingivitis (308). Some cells do penetrate the oral mucosa in healthy subjects, presumably as a result of diapedesis (307). Loss of leukocytes in the urine also has been demonstrated in normal subjects (309). Leukocyte loss increases significantly in association with pyelonephritis (310). In addition, arteriovenous catheterization studies in dogs (311) and in humans (312) provide evidence that suggests leukocytes also are removed in the lungs, liver, and spleen. Significant numbers may be lost into the gastrointestinal tract. No quantitative data concerning the rate of loss through these various organs are available. Interestingly, bone marrow and blood leukocyte counts were essentially the same in germ-free and normal mice, suggesting that loss via subclinical infections at the various body surfaces is not a major factor in neutrophil turnover (313,314).
Neutrophil Kinetics in the Fetus and Newborn
The fetus exists in a sterile environment and, unlike the adult, does not require antibacterial defenses. However, if the systems ensuring adequate neutrophil production, storage, delivery to tissues, phagocytosis, and bacterial killing are not intact at birth, extrauterine existence is seriously compromised. Maturation of the neutrophil system is not complete in the midgestation fetus, and even the neonate delivered at term has a neutrophil system that in several respects is quantitatively and qualitatively immature. Therefore, one might anticipate that newborns, particularly those delivered prematurely, would be at significant risk for serious bacterial infection, and many studies reveal a strong correlation between prematurity and serious bacterial infection (315,316,317,318,319,320). Of all the risk factors for bacterial infection analyzed by the national collaborative study on neonatal infection, premature birth showed the strongest correlation (321). In addition, babies delivered at term experience a higher incidence of bacterial infection than do older children or adults (321,322) and, when infected, often display poor resolution of infection despite antibiotic therapy.
Newborn infants, particularly if premature, display many other deficiencies in antibacterial defense, such as low levels of IgG antibody, complement components, fibronectin, and lymphokine production (323), but only maturational differences in neutrophil kinetics are discussed here. Realizing that the neutrophil system of a fetus is underdeveloped and in a state of maturation, a difference in neutrophil pool sizes and kinetics in this group from those in adults can be expected. In addition, rapid somatic growth in the fetus and newborn places added demands, unique to the neonate, on neutrophil production (324); cells are needed not only for ongoing antibacterial defense, but also for a rapidly increasing body mass.
The investigation of neutrophil kinetics during fetal and neonatal life in humans has been hampered by lack of applicability of the techniques used for such studies in adults. For instance, DF32P and 3H-TdR blood kinetic measurements have not been used in babies because of the radiation exposure and the large volumes of blood required. Nevertheless, the results of clinical studies, coupled with extensive investigation in developing animals, illustrate several developmental differences.
Granulocyte-Macrophage Progenitor Cells
CFU-GM has been isolated from the livers of aborted 5- to 6-week human fetuses (325). Although early fetal CFU-GM produced colonies of mature neutrophils in vitro, no mature neutrophils could be located in the liver, marrow, or blood of the 5- to 6-week fetuses from which this CFU-GM was obtained (325). In separate studies, mature neutrophils were not detected in the human fetus until approximately 15 to 16 weeks’ gestation (326,327,328,329), at least 10 weeks after the first appearance of CFU-GM.
The concentration of CFU-GM in blood is higher in the fetus and newborn than in adults: 20 to 300 CFU-GM/ml of venous blood in adults and approximately 2000 CFU-GM/ml in term neonates (330,331,332,333,334,335,336,337). Even higher venous blood concentrations are found in prematurely delivered infants (331,336), with the highest values noted in the most premature subjects (331). The total body pool of CFU-GM has not been measured in human neonates; in rats, the number of CFU-GM/g body weight is small in the fetus (0.5 ± 0.1 × 103 CFU-GM/g) and increases to adult levels (10.5 ± 0.2 × 103/g) at 4 weeks of age (338).
The proliferative rate of CFU-GM during human gestation, assessed by thymidine suicide, is rapid in the second trimester (331). Whereas CFU-GM in venous blood of adults displays a thymidine suicide rate ranging from 0 to 7% (331,339,340,341), rates of 45% have been observed in term neonates, and rates of 75% were noted in prematurely born neonates (331). Similarly, in fetal and neonatal rats (243) and mice (340), the CFU-GM proliferative rate is high and appears to be near maximal at birth, even in the noninfected state (342,343). Unlike in adult animals, no further increase in CFU-GM number (344) or CFU-GM proliferative rate (342) has been detected during either sublethal or lethal bacterial infection, suggesting that the system operates at capacity. It has been suggested that the inability to accelerate neutrophil production above baseline might handicap neonates by limiting their neutrophil supply during a bacterial infection.
Neutrophil Storage Pool
In the fetus, as in the adult, mature neutrophils are stored within the skeletal marrow but also are found in the liver and spleen (344). Techniques that measure the size of the neutrophil storage pool, such as radioisotopic iron labeling, with subsequent liver, spleen, and bone marrow biopsy, have not been applied to normal human neonates. In fetal and neonatal animals, however, the liver and spleen, as well as the long bones, can be removed, and the neutrophils within them can be quantified. Such studies in rats show that the neutrophil storage pool is considerably smaller in prematurely delivered animals (1.0 to 1.3 × 106 cells/g body weight) than in term (1.3 to 2.5 × 106 cells/g) and adult animals (4.5 to 7.5 × 106/g) (345).
During experimental bacterial sepsis in neonatal dogs and rats, the size of the neutrophil storage pool has been serially quantified. Experiments performed with a variety of organisms (344,346,347,348,349,350,351,352) demonstrate depletion of the storage pool and neutropenia before death. Similarly, in human neonates with lethal bacterial sepsis, neutropenia and depletion of the neutrophil storage pool, as assessed by bone marrow aspiration, are nearly universal findings (328,350,353,354).
Release from Storage Pool into Blood
Within 1 hour of inoculating adult animals with as few as 104 type III group B streptococci/g body weight, an accelerated rate of egress of neutrophils from the storage pool into the blood can be detected (353). In contrast, when newborn animals were given the same or a larger inoculum of 106 organisms/g, a 4-hour delay occurred before this acceleration in neutrophil storage pool release was observed (353). This delay between bacterial inoculation and accelerated release of marrow neutrophils may be a significant physiologic disadvantage, which permits prolific initial bacterial multiplication. In other studies, this delay in release of marrow neutrophils was completely corrected by prior administration of type-specific antibody directed toward the organism with which the animal was inoculated (347,353). Some cytokines (e.g., IL-8) readily induce neutrophil release from the marrow (361).
Alterations in Migration into Tissues and Sites of Destruction in Neonates
In many studies, investigators have demonstrated defective neutrophil chemotaxis in neonates. In early investigations, Eitzman and Smith, using the Rebuck skin window technique, demonstrated that a preponderance of eosinophils, not neutrophils, was attracted to the abraded dermis of neonates (355). Using the same technique, Bullock et al. demonstrated that neutrophils in neonates remained at the site of abrasion longer than they did in adults (356).
Using the Boyden chamber method, neutrophils from newborns were found to be less responsive than adult neutrophils in chemotaxis (357). In addition, factors generated from neonatal serum attracted neutrophils less well than did factors from adult serum. Diminished chemotaxis of cord blood neutrophils was also demonstrated, with reduction to approximately 80% of levels observed with adult neutrophils (358,359,360). Using a different in vitro technique for quantification of neutrophil movement (agarose gel), newborn neutrophil chemotaxis was found to be reduced to approximately 20 to 27% of that seen with adult neutrophils (361). In prematurely delivered neonates, neutrophil chemotaxis was even more defective (362), and the defect persists for a considerable time after birth.
A further reduction in chemotaxis of neutrophils from ill neonates compared to healthy neonates (363) and decreased chemotaxis in preterm neonates with bacterial sepsis followed by a return to normal neonatal values (approx. 20% of adult values) with resolution of the infection (364) have been reported.
Neutrophil migration has also been investigated in vivo in neonatal and adult animals. One technique involved surgical implantation of a sterile polyvinyl sponge disc, standardized for body weight, into rats. At various intervals, the sponges were removed, and neutrophils that had migrated into them were chemically quantified. Concurrent with sponge removal, the long bones (and the liver and spleen in neonates) were also removed, and the size of the total body neutrophil storage pool was determined (349). By comparison with control animals, only approximately 9% of the neutrophils released from the storage pool in neonates could be accounted for in the sponge. In contrast, approximately 60% of the neutrophils released from the marrow in adults migrated to the sponge, indicating more efficient neutrophil migration in the adult. Another study found that the accumulation of neutrophils in the peritoneal cavity of rats after intraperitoneal inoculation with various chemical attractants was impaired in neonates (365).
The mechanism responsible for deficient neutrophil chemotaxis in neonates is only partly known. By measuring the pressure needed to aspirate neutrophils into a glass pipet, Miller determined that the neonatal neutrophil was more rigid and less deformable than the adult neutrophil, a characteristic that may be detrimental to movement of neutrophils through tissues (366). In addition, neutrophils from human neonates irreversibly aggregated after activation by C5a, whereas after exposure to the same stimulus, adult neutrophils aggregated and then deaggregated (265,266,267,268,367). Irreversible aggregation of neonatal neutrophils has also been seen in response to activation by FMLP (368). If irreversible neutrophil aggregation occurs in vivo in neonates, then independent neutrophil mobility would probably be impaired. A deficiency in the redistribution of cell-surface adhesion sites, factors related to impaired migration, and impairment of neutrophil adhesiveness has also been reported (369,370) and would impair neutrophil function in vivo. Response to complement-derived peptides was also impaired, as shown in the response of cord blood neutrophils to endotoxin-activated serum (371). Concanavalin A capping (372), phytohemagglutinin-induced aggregation (373), and C5a-induced chemotaxis (348), as well as FMLP-induced membrane potential change (374), are reduced in the neonatal neutrophil. No difference in calcium uptake in FMLP-stimulated neutrophils was noted between human neonates and adults, but reduction in calcium uptake by resting neonatal neutrophils was significant (375). Diminished orientation of neonatal neutrophils after exposure to a chemotactic gradient was noted, and a significant decrease both in the number of microtubules/cell and in the assembly of microtubules was observed (376).
![]() Physiologic Variation in Leukocytes
Physiologic Variation in Leukocytes
The changes in blood leukocyte concentration that occur with growth and development are shown in Appendix A. By the age of 4 to 8 years, the blood differential cell count approaches that seen in the adult. Normal values are presented in Table 9.6. Metamyelocytes or myelocytes are not often seen on routine examination of the blood smear, but a few such cells can be found in normal blood after a careful search or, more readily, by examination of buffy coat smears (3.6/3,000 granulocytes) (377); atypical mononuclear forms and megakaryocyte fragments containing nuclei are also seen in such smears. Whether a significant difference exists in leukocyte concentration between the sexes or with advancing years has been debated (165,378,379,380,381). Racial variations were reported in black Africans, with lower neutrophil and monocyte counts and higher eosinophil counts; however, these differences were not as evident in Africans eating a Western diet (382) or in black Americans (129,180).
Although leukocyte concentration is maintained within definite limits in normal humans, fluctuations occur during a single day and from day to day. The suggestion that a characteristic hourly rhythm occurs has not been confirmed (383,384), nor has the occurrence of a digestive leukocytosis been established (385). Light influences the diurnal variation of neutrophils (220,386). Under conditions of complete physical and mental relaxation, a basal level of 5.0 to 7.0 × 109 cells/L is usual (387). Ordinary activity may be associated with a moderate increase, and a somewhat higher level is common in the afternoon. Under all these conditions, however, the leukocyte count tends to remain within the normal range (Table 9.6).
Conclusive demonstration of the effects of climate or season on the leukocyte count is lacking. Some authors claim that meteorologically conditioned fluctuations occur (388). Heat and intense solar radiation are said to cause leukocytosis (389). Prolonged residence in Antarctica has been reported to cause leukopenia (390). Artificially induced heat, sunlight, and ultraviolet light have been reported to cause lymphocytosis (391). Acute anoxia, both anoxic and anemic, causes neutrophilic leukocytosis (392) that does not develop in adrenodemedullated rats. In the first few days after a subject has arrived at a high altitude, some leukocytosis, accompanied by lymphopenia and eosinopenia, has been observed, followed quickly by slight lymphocytosis and eosinophilia (393).
Marked leukocytosis occurs regularly with strenuous exercise. Counts as high as 22.0 × 109/L have been recorded for a runner after an 11-second 100-yard dash, and 35.0 × 109/L has been recorded after completing a quarter-mile run in less than 1 minute (387). The increment of cells usually consists of segmented neutrophils, but lymphocytosis may be prominent as well. Such leukocytosis recedes to normal in less than 1 hour and, in the neutrophil series, is related to a shift of cells from marginal sites (MGP) to the circulation (CGP) (shift leukocytosis) (258,259). This leukocytosis occurs in the absence of the spleen, suggesting that the spleen is not a major site of cell margination. Leukocyte counts >20.0 × 109/L, mainly neutrophils, are regularly recorded for runners who complete a 26-mile marathon in 2.5 to 3.0 hours; whether a shift to the left, suggesting mobilization of marrow neutrophils, occurs in this circumstance is debatable (387). Postmarathon leukocytosis subsides slowly over a number of hours and probably reflects a redistribution of granulocytes in the blood, combined with mobilization of cells from the marrow with an increase in TBGP size. The magnitude of the leukocytosis associated with exercise appears to depend primarily on the intensity of the activity rather than on its duration (394).
Convulsive seizures, from whatever cause, are associated with an increase in leukocyte count similar to that noted after violent exercise. Electrically induced convulsions are followed by a reduction in eosinophil and lymphocyte number and an increase in neutrophil number, findings consistent with the effects of adrenal hormone secretion (395). Epinephrine injection produces leukocytosis, the nature and duration of which appear to vary with the mode of administration. Intramuscular injection causes leukocytosis in two phases (396,397). In the first phase, maximal at 17 minutes, the number of neutrophils, lymphocytes, and eosinophils increases and then returns toward normal over several hours. This pattern almost certainly represents a shift leukocytosis. In the second phase, the number of neutrophils rises again at approximately 4 hours, although the number of lymphocytes and eosinophils remains at or below preinjection levels (397); this phase may reflect an adrenal steroid effect and consists of an absolute neutrophilia. After intravenous injection, leukocytosis peaking at 5 to 10 minutes and of total duration of less than 20 minutes occurs and has been shown to be purely shift neutrophilia (259,260). The leukocytosis that follows subcutaneous injection is more variable.
During attacks of paroxysmal tachycardia, leukocytosis with cell counts of 13.0 to 22.0 × 109 cells/L has been reported (398). Pain, nausea and vomiting, and anxiety may cause leukocytosis in the absence of infection (399). The paucity of band forms and metamyelocytes indicates that the neutrophilia results from the redistribution of the cells between the marginal and circulating pools.
Ether anesthesia produces leukocytosis, probably because of emotional and reflex reactions, as well as struggling during the stage of excitement. Narcosis with barbital compounds usually reduces the leukocyte count.
During the ovulatory period, eosinopenia and a slight rise in the number of leukocytes, as well as increased 17-hydroxycorticosteroid levels, have been reported (400,401). Slight leukocytosis occurs during pregnancy, and neutrophilia increases as term approaches (180,400). The onset of labor is accompanied by neutrophilic leukocytosis, which sometimes is pronounced (34.0 × 109/L). This state continues for 1 day after delivery, receding to normal only after 4 or 5 days. These changes are accompanied by a reduction in the number of circulating eosinophils (400).
Many of the physiologic variations in leukocytes that have been described can be explained as manifestations of stimulation of the adrenal cortex. The administration of cortisone or hydrocortisone results in increased blood levels of 17-hydroxycorticosteroids that peak at 1 hour (402) and are associated with neutrophilia (403). Eosinopenia and lymphopenia follow, become maximal at 4 to 8 hours, and are proportional to the quantity of hormone administered. Neutrophilia was less constant than the depression in eosinophil and lymphocyte numbers but is probably caused by a steroid hormone–mediated decreased efflux of neutrophils from the blood and increased cell release from the bone marrow (260,262). Just as exercise can raise the circulating neutrophil count, intense exercise has also been reported to induce neutrophil activation as determined by studies of cell-surface antigen expression (404). In contrast, ultraviolet B radiation, at doses similar to those naturally received, inhibits neutrophil phagocytosis and adhesion, although the practical significance of these observations is unknown (405).
![]() Control Mechanisms Regulating Neutrophil Production
Control Mechanisms Regulating Neutrophil Production
It is evident that a true steady state of neutrophil kinetics exists only for brief periods. Shifts of cells between marginal and circulating sites may occur without changes in blood neutrophil turnover (260), but any change in TBGP size must result from changes in cell inflow or egress. Studies involving leukapheresis have shown that a normal animal replenishes a depleted TBGP by mobilizing cells from the marrow granulocyte reserves (274). This increase in neutrophil concentration and TBGP size, like that seen with most bacterial infections or after endotoxin or steroid administration, must be triggered by some signal, and some means of stimulating cell production must be available to replenish depleted marrow reserves, whatever the etiology.
The nature of these control mechanisms is not well understood, but several control points exist: recruitment of pluripotent stem cells and their induction into committed stem cells, stimulation (and perhaps inhibition) of stem cell and myeloid proliferative cell growth, and selective release of cells from the marrow.
Blood cell development is discussed in Chapter 5 and is only briefly discussed here. Pluripotent stem cells are mostly in the G0 state and must be induced into actively proliferating committed stem cells. Hematopoietic cell growth and development are usually restricted to certain tissues (e.g., bone marrow in adult humans and bone marrow and spleen in mice). Because cell differentiation is influenced by organ microenvironment (e.g., erythropoiesis is favored in mouse spleen, but granulocytopoiesis is favored in the bone marrow), the concept of local control of pluripotent stem cell induction was developed (the hematopoietic microenvironment) (406). The importance of the hematopoietic microenvironment is exemplified by the anemia of Steel mice, which results from a defective hematopoietic microenvironment (406). The defect in Steel mice involves the granulocyte system as well as erythropoiesis (407) and results from a diminished ability of organ stroma in the bone marrow and spleen to induce committed stem cells from pluripotent stem cells (408). The demonstration that the Steel (Sl) gene product, deficient in Steel mice, is a growth factor (stem cell factor or stem cell colony-stimulating factor) that binds to a receptor coded by the c-kit proto-oncogene (or white-spotting locus, W) has provided a molecular basis for understanding the hematopoietic abnormalities in mice with genetic defects in the Steel or W genes (256,409).
As judged from suspension cultures, at least three cell types (giant fat cells, epithelial cells, and phagocytic mononuclear cells) provide the microenvironment needed for multipotent stem cell proliferation (410). Presumably, these stromal cells produce sufficient concentrations of hematopoietic cell growth factors locally to promote multipotent stem cell proliferation and renewal when required (411).
Growth Factors
A large number of growth factors or colony-stimulating factors have been identified that regulate neutrophil production in the bone marrow, as described in Chapter 5. Two of these, G-CSF and GM-CSF, are in clinical use. Exogenous administration of G-CSF expands the granulocyte mitotic pool and also decreases the bone marrow transit time of the postmitotic cells without changing the blood neutrophil half-life (412). These factors not only speed the recovery of neutrophil counts after chemotherapy and may decrease associated infectious complications (413,414,415,416,417), but also have effects on mature neutrophils. For example, G-CSF transiently increases CD11b expression (414,418,419) and the affinity of CD62L (L-selectin) for its ligand and then causes CD62L surface expression to decrease (420). G-CSF also primes neutrophils for subsequent superoxide production in response to FMLP (421,422,423,424). Intravenous administration of G-CSF can result in an immediate transient neutropenia (425,426) similar to the transient increase in the MGP seen with hemodialysis. Evidence of neutrophil degranulation in vivo after administration of G-CSF has also been observed (427). A number of other cytokines among this rapidly expanding class can also activate or prime neutrophils, including TNF-α, IL-6, IL-1, and IL-8 (428,429,430,431).
In addition to locally produced stimulators of colony-forming unit stem cell proliferation (432), inhibitors of proliferation have also been described (433,434). Lactoferrin (present in the secondary or specific neutrophil granule) binds to specific receptors on some monocyte-macrophages and suppresses release of GM-CSF (and other cytokines), thus inhibiting colony formation. Transferrin also exhibits colony-suppression activity, possibly through inhibition of GM-CSF production by T lymphocytes (435). Soluble forms of receptors for cytokines may also regulate the response of bone marrow progenitors to growth factors (436). Neural mechanisms controlling hematopoietic cell proliferation and release have also been suggested (437).
Another control point in the system is the selective release of granulocytes from the marrow. In studies of perfused rat hind limbs, the release of neutrophils from the marrow into the blood (438) increased with an increase in perfusion flow rate or with a low leukocyte content of the perfusate (439). Serum or plasma from animals or humans made neutropenic by endotoxin, vinblastine, or nitrogen mustard also induced neutrophilia (440,441,442,443). The activity was present during the period of neutropenia and rising neutrophil concentration—but not before or after this period. This neutrophilia-inducing activity was qualitatively dissimilar from that noted after endotoxin, epinephrine, or cortisone administration and acted by causing release of marrow cells. The results of these studies suggest that an endogenously produced humoral factor causes neutrophil release from the marrow (440). Several factors that stimulate neutrophil release from the bone marrow have been identified, including G-CSF, GM-CSF, C5a, TNF-α, TNF-β, IL-8, and, possibly, a cleavage product of the third component of complement (203,257,258). Studies in rabbits have found that IL-8 induces neutrophil release from the bone marrow without altering the transit time through the mitotic and postmitotic marrow pools (258).
Thus, evidence exists that several factors, including cell nuclear and cytoplasmic deformability, cell motility, affinity of cell adhesion molecules for ligands, blood flow, and others are important in the control of marrow cell release (103,104,256,444).
![]() Neutrophil Function
Neutrophil Function
The major role of neutrophils is to protect the host against infectious agents. To accomplish this task, the neutrophil must first sense infection, migrate to the site of the infecting organism, and then destroy the infectious agents. Although neutrophils can sense a stimulus in suspension, they can migrate only when in contact with a surface. Thus, although in some cases neutrophils in blood may respond to a stimulus by adhering to other blood cells or foreign bodies, such as bacteria or biomaterials, the usual first step of the neutrophil after sensing an inflammatory stimulus is to adhere more strongly to the blood vessel wall. Usually, this occurs in a postcapillary venule. After adhesion to the endothelial surface, the neutrophil follows a gradient of chemotactic factors to the site of infection and interacts with the organisms. Finally, when the neutrophil reaches the infecting organism, it must destroy it. This destruction is generally accomplished by phagocytosis of the agent followed by release of granules into the phagocytic vesicle, followed by killing of the organism. The mechanisms by which these phenomena occur are very complex and not completely understood.
History
Antibacterial properties of blood were described by the British surgeon John Hunter in ∼1761 during the Seven Years War. He observed that the cellular (buffy coat) component of blood could retard the “spoilage” of blood (445); we now would recognize these effects as reflecting the antibacterial properties of leukocytes. In these classic studies, Hunter observed that when blood was allowed to stand, a “buff colored” layer was visible on top of the red cells. He noted that blood from patients with infected wounds had a thicker “buff colored inflammatory crust” than observed in blood from healthy subjects, which we now understand reflects the neutrophilia associated with infection. With time, he noted that blood would “spoil,” as determined by the development of an odor typical of spoiled food. Hunter found that the addition of the “buff colored inflammatory crust” to a blood sample would delay the time to “spoilage,” now understood to reflect the antibacterial abilities of neutrophils.
The response to infection by neutrophils in the microvasculature was elegantly described in An American Text-Book of Surgery in 1892 (297). This description remains instructive and is summarized here. At one time, the inflammatory cells at sites of infection were thought to be caused by proliferation of connective tissue cells. After the observations of von Recklinghausen that many of these cells were capable of locomotion (and called “ameboid cells from their resemblance to the amoeba”), Cohnheim identified the cells in the inflamed tissue as leukocytes (297). Microscopic examination of a frog’s mesentery or tongue reveals “an arteriole with its rapid pulsating current of blood, and near by a small vein in which the blood flows with a more steady movement. The red blood-corpuscles occupy the axis of the blood vessel, and the few white corpuscles which are seen float in the more sluggish stream of plasma which occupies the borders of the lumen and appears as a transparent layer” (Fig. 9.14). Induction of inflammation by the application of a caustic agent leads to hyperemia. “The rapidity of the flow of blood is greatly increased and a greater amount of blood is observed in the part. The lumen of the artery is greater than before, and the column of red corpuscles is much broader, and fills a comparatively greater portion of the lumen of the vessel. The capillaries are now quite distinctly seen, and are crowded with blood-corpuscles” (Fig. 9.15). This is followed by
a slowing of the current which soon becomes much more sluggish than in the normal state. This is first noticed in the capillaries, and soon after in the veins. The pulsation, however, continues in the arteries. As a result of this diminution of speed the column of blood-corpuscles becomes broader and almost completely fills the interior of the vessels. In the veins a great accumulation of white corpuscles takes place on the interior of the walls—. Finally, they are so greatly increased in numbers that the entire wall of the vessel appears to be lined with leukocytes. The white corpuscles also accumulate in the capillaries, but not to the same extent [Fig. 9.16].
|
|
|
Figure 9.14. Diagram of normal vessels and bloodstream: a, artery; b, vein; c, capillary. (From Keen WW, White JW, eds. An American textbook of surgery. Philadelphia: WB Saunders, 1892;11, with permission.) |
The margination of the neutrophils is shown schematically in Figure 9.17 (platelets or “blood-plaques” are also shown).
|
|
|
Figure 9.15. Diagram of dilation of the vessels in inflammation: a, artery; b, vein; c, capillary. (From Keen WW, White JW, eds. An American textbook of surgery. Philadelphia: WB Saunders, 1892;11, with permission.) |
|
|
|
Figure 9.16. Diagram of stasis of blood and diapedesis of white corpuscles in inflammation: a, artery; b, vein; c, capillary. (From Keen WW, White JW, eds. An American textbook of surgery. Philadelphia: WB Saunders, 1892;12, with permission.) |
|
|
|
Figure 9.17. Blood plaques or third corpuscles (a). Red corpuscles (b). White corpuscles (c). (From Keen WW, White JW, eds. An American textbook of surgery. Philadelphia: WB Saunders, 1892;14, with permission.) |
Beginning concurrently with the slowing of the blood-stream, is the emigration of the leukocytes from the interior of the veins. Many leukocytes, by a change of shape, send out little prolongations of protoplasm into the substance of the wall, and slight protuberances are soon seen projecting from its outer surface. These enlarge, and we now see the corpuscles presenting an hour-glass appearance. The portions within the vessel soon follow those without, and the leukocytes escape from all contact with the vessel. Many corpuscles appear to follow one another through the same point in the wall [Fig. 9.18].
Migration takes place to a limited extent also from the capillary vessels, but no such process is observed in the walls of the arteries. These same actions of neutrophils, as well as the formation of neutrophil-neutrophil aggregates with infections in rabbit ear veins, were videotaped by W. B. Wood in the 1960s. Using fluorescein-labeled neutrophils, Hammerschmidt et al. later demonstrated that injection of C5a reproduced the increase in neutrophil margination and aggregation in rat mesentery (446).
Early views of neutrophil function at the site of inflammation included a role in the repair process. Later, their ability to phagocytose was recognized. Metchnikoff advanced the theory known as phagocytosis, according to which the cells of the inflamed part, by virtue of their ability to consume foreign substances, attack and destroy the invading bacteria (Fig. 9.19).
These cells are called phagocytes (from the Greek, “to eat,” and, “cell”). If they are able to destroy the bacteria, the system is protected from the invading organisms. The leukocytes are called micro-phagocytes (or microphages), and the larger cells developed from the fixed connective-tissue cells are called the macro-phagocytes (or macrophages) (297).
|
|
|
Figure 9.18. Diagram of stages of the migration of a single white blood corpuscle through the wall of a vein in 1 hour and 50 minutes (mesentery of the frog).(From Keen WW, White JW, eds. An American textbook of surgery. Philadelphia: WB Saunders, 1892;13, with permission.) |
|
|
|
Figure 9.19. Diagram of a phagocyte destroying a bacillus. The cell is shown at three different times (a, b, and c). (From Keen WW, White JW, eds. An American textbook of surgery. Philadelphia: WB Saunders, 1892;16, with permission.) |
Chemotactic Factor Receptors
The initial step of the neutrophil response to infection is the detection of an appropriate signal. The interaction of bacteria with blood components, especially antibodies and the complement system, results in the formation of various chemotactic factors. In some instances, the bacteria directly release factors that are chemotactic for neutrophils. The interaction of bacteria or their products with other host cells may also result in the formation of chemotactic factors. Neutrophils express specific receptors on their cell surface for a variety of chemotactic factors. These receptors include those for n-formylated peptides such as n-formyl-met-leu-phe (FMLP), C5a, leukotriene B4, and platelet-activating factor. The initial activation of the neutrophil occurs when soluble chemotactic factors bind their receptors on the neutrophil surface. As with the interaction of antibodies and haptens, the association kinetics for these receptor–ligand interactions are very rapid. Typically, sufficient receptor–ligand interaction to initiate neutrophil activation occurs within seconds.
Signaling
Many chemotactic factor receptors appear to be coupled to guanine nucleotide-binding proteins (G proteins) (447). The role of G proteins in neutrophil signal transduction is supported by a variety of studies, including classic studies using pertussis toxin. Pertussis toxin adenosine diphosphate ribosylates certain G proteins and inhibits neutrophil responses to a number of stimuli, including FMLP. In contrast, some neutrophil responses, such as phorbol ester–induced secretion, are not inhibited by pertussis toxin. Although the details of signal transduction in neutrophils are not fully elucidated, it appears that the G proteins associated with chemotactic factor receptors are important in receptor activation of phospholipase C, which then hydrolyzes phosphatidylinositol bisphosphate (PIP2), resulting in the generation of two second messengers, IP3 and 1,2-diacylglycerol (DAG). Experiments suggest that IP3 binds to specific receptors on intracellular membranes, resulting in the release of calcium from intracellular stores, which is rapidly augmented by an influx of extracellular calcium. Thus, shortly after receptor–ligand binding, the intracellular calcium rapidly rises from a resting level of approximately 0.1 mmol/L to approximately 1 mmol/L (448,449,450). This rise in free intracellular calcium is transient and returns to baseline in approximately 1 to 3 minutes.
It appears that the initial rise in intracellular calcium caused by the release of intracellular calcium stores plays a critical part in the alteration of membrane permeability to allow the influx of extracellular calcium. To some extent, variations in intracellular calcium transients may direct specific cellular functions, in that specific granule release occurs at very low (submicromolar) free calcium concentrations, whereas in studies using permeabilized cells, higher (micromolar) levels of free calcium result in release of both specific and azurophil granules (451,452,453). Although the extracellular calcium influx is critical for many neutrophil responses, it is not critical for all, as degranulation is not blocked by ethyleneglycol-bis(2-aminoethyl) tetra-acetic acid (454). Similarly, phagocytosis of particles opsonized with C3bi can occur without apparent intracellular calcium transients (455).
Protein phosphorylation is an important mechanism of the regulation of protein function, and a number of studies suggest its role in neutrophil activation. Phorbol myristate acetate, which activates a number of neutrophil functions, binds to PKC and results in its activation. Although phorbol myristate acetate is not present in neutrophils, DAG, released when phospholipase C hydrolyses PIP2, also binds and activates PKC. Because calcium is also important in PKC activation and IP3 increases intracellular calcium, the hydrolysis of PIP2 to IP3 and DAG may contribute to PKC activation via both DAG and IP3. Interestingly, the specific granules have been reported to contain a PKC inhibitor, thus providing a possible mechanism to down-regulate PKC-mediated responses (456).
Although this model explains many observations, it has become clear that signal transduction in neutrophils is far more complex, with both Ca2+and PKC-independent pathways. Tyrosine phosphorylation has been found to play a critical role in signal transduction from various chemotactic factor receptors. Multiple neutrophil proteins are rapidly phosphorylated after activation, including Src family kinases; the Lyn kinase is activated by chemotaxins, increasing its ability to phosphorylate substrates. Serine and threonine kinases also appear to be involved in signaling, and some are activated by FMLP. In addition, phosphatidylinositol 3-kinase is also activated by chemotaxins. Phosphatidylinositol 3-kinase catalyzes the phosphorylation of PIP2 to PIP3. Phosphatidylinositol 3-kinase binds some phosphotyrosine residues via the SH2 domain on one of its subunits. Protein tyrosine phosphatases probably also play a role, as the transmembrane protein phosphatase CD45 has been implicated as a regulator of neutrophil function (47,457). Chemotactic factors have also been found to activate phospholipase A2 and phospholipase D. Finally, the importance of low-molecular-weight guanosine triphosphatases (LMWG) is also being recognized. Knowledge of signal transduction is rapidly advancing, and the reader is referred to reviews (458,459) and the current literature.
Physiologic soluble inhibitors of neutrophil function have also been identified. For example, adenosine inhibits neutrophil aggregation, adhesion, chemotaxis, and superoxide production (460,461,462,463). These inhibitory effects appear to act via A2 receptors without preventing the transient rise in intracellular Ca2+ (460,461,462).
Neutrophil Priming
“Priming” is an important concept in neutrophil signaling. Signaling in neutrophils is complex and can be initiated by many different stimuli that may share downstream signaling pathways. When neutrophils are exposed to an appropriate low level of a stimulus, they can be primed to a condition such that they display a much more prominent response to a second stimulus than they would if they had not been primed (463,464,465,466). Neutrophils can be primed by one stimulus for a response to a different agonist. Priming occurs at doses that do not result in a rise in cytoplasmic free calcium, but still cause protein tyrosine phosphorylation of signaling molecules. After priming, neutrophils exhibit a more prominent respiratory burst or secretory response to a given stimulus than would occur if priming had not occurred. This phenomenon may be involved in many physiologic neutrophil responses in vivo. Neutrophil priming can be reversible, with the cells still capable of being reprimed (467).
Desensitization
After previous exposure to a stimulus, neutrophils react less to subsequent stimulation by the same stimulus (468,469,470). This phenomenon has been termed desensitization. In some cases, the desensitization appears to be specific to the original stimulus, but in other cases desensitization to different stimuli is also observed (cross-desensitization) (468,469,470,471).
Such desensitization has been observed in patients undergoing hemodialysis, in which exposure of blood to a cuprophane dialyzer membrane results in the generation of C5a, which causes a transient neutropenia due to pulmonary leukostasis, as described in Chapter 9 (265,266). Although C5a generation persists throughout dialysis, the neutropenia is transient (267). In contrast to neutrophils obtained at the start of dialysis, neutrophils obtained after 2 hours of dialysis (after the leukostasis has resolved) do not aggregate in response to plasma leaving the dialyzer membrane, demonstrating desensitization in vivo (267). A patient with cytomegalovirus infection, whose serum induced granulocyte aggregation (presumably due to C5a), did not experience neutropenia during dialysis, and his neutrophils did not aggregate in response to serum leaving the dialyzer, in contrast to control cells, also demonstrating in vivo desensitization (267). Similar desensitization was demonstrated in rabbits using the chemotactic peptide FMLP, wherein continuous intravenous infusion of FMLP reproduced a transient neutropenia due to pulmonary sequestration (472). It is likely that neutrophil desensitization may also occur in other pathologic states, including infection, trauma, and multiorgan failure syndrome, and may contribute to neutrophil dysfunction, although the clinical significance of this phenomenon is unclear.
Neutrophil function can also be regulated by the neuroendocrine axis. For example, glucocorticoids can regulate neutrophil function, at least partly through annexin I or lipocortin I (75). The phenomenon of desensitization has also been implicated as the mechanism by which glucocorticoids inhibit neutrophil function. It appears that the glucocorticoid-regulated protein annexin 1 (lipocortin 1) can bind a formyl-peptide receptor, induce calcium transients, and desensitize neutrophils to subsequent stimulation by other agents (471). Two formyl-peptide receptors are expressed in neutrophils, the classical FPR and the related receptor FPRL-1/AXL. Annexin 1 appears to bind to FRPL-1/AXL, whereas some peptides derived from annexin 1 may bind both receptors (474). Opiates have been reported to alter many immune functions, including neutrophil functions such as the respiratory burst, in many species (473). MRP-14, a prominent component of neutrophils, can inhibit the function of activated macrophages and, possibly by this mechanism, can decrease inflammatory pain (74).
Neutrophil–Endothelial Adhesion
Both neutrophils and endothelial cells express a variety of adhesive molecules on their cell surface, and the expression and activity of these molecules in many cases can be regulated by stimuli. Some of the known adhesion molecules of neutrophils and endothelial cells are indicated in Table 9.8. Approximately one-half of the circulating neutrophils exist in the so-called marginating pool, some of which can be seen microscopically to be rolling along the endothelial surface, maintaining a loose intermittent contact with endothelial cells. The importance of hemodynamic forces, especially of red cells, in directing leukocytes outward from the flowing blood toward the endothelium was described many years ago and subsequently confirmed (297,476,477,478). An attractive model of neutrophil–endothelial cell adhesion has been proposed by Springer, which accounts well for the known data (479). In this model, selectin molecules on the cell surface are responsible for neutrophil rolling along the vessel wall. This loose adhesion brings the neutrophil in close proximity to the endothelial cell, where chemoattractants can be released or displayed on the cell surface. The interaction of these chemoattractants with neutrophil receptors results in signal transmission and the activation of integrin molecules. These integrins can then bind their ligands on the endothelial cell surface, resulting in a marked increase in adhesion to the endothelial cell and cessation of rolling. After this, the cells sense further chemoattractant gradients and migrate into the tissue.
|
Table 9.8 Neutrophil–Endothelial Cell Adhesion Proteins |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Selectins
Three selectins have been identified that each have an N-terminal domain that is homologous to Ca2+-dependent lectins (Fig. 9.20). L-selectin (CD62L, LAM-1, LECAM-1) is expressed on the neutrophil surface. The ligand for L-selectin is the glycoprotein known as Gly-CAM-1(glycosylation-dependent cell adhesion molecule-1). Endothelial cells express both E-selectin (CD62E, endothelial-leukocyte adhesion molecule-1) and P-selectin (CD62P, granule membrane protein-140, platelet activation–dependent granule-external membrane protein). E-selectin and P-selectin both recognize Lewisx-related sialylated carbohydrates. Expression of E-selectin on the endothelial cell surface can be induced with stimuli such as IL-1 and TNF but requires protein synthesis. In contrast, P-selectin (CD62P) is found in both the Weibel-Palade granules of endothelial cells and the platelet α-granule. Thus, stimulation of endothelial cells with the appropriate stimulus, such as thrombin or histamine, can result in a rapid mobilization of CD62P (P-selectin) to the endothelial cell surface.
Integrins
Integrins are noncovalently associated heterodimers of α and β subunits, each of which has characteristic structural motifs (Figs. 9.21 and 9.22). The major integrins of neutrophils are the β2 integrins made up of αLβ2 (leukocyte function antigen-1, CD11a/CD18), αmβ2 (HMac-1, CD11b/CD18), and αxβ2 (p150,95, CD11c/CD18). Intercellular adhesion molecule (ICAM)-1 (CD54) expressed on the endothelial cell surface is a ligand for both CD11a/CD18 and CD11b/CD18. Other Ig superfamily members are probably also involved in neutrophil–endothelial cell adhesion, including platelet endothelial cell adhesion molecule-1 (CD31), ICAM-3 (CD50) (expressed on the neutrophil but not the endothelial cell), and the CD66 family of neutrophil-activation antigens.
|
|
|
Figure 9.20. Schematic of selectin structure. The selectins are attached to the cell via a transmembrane domain with an extracellular domain consisting of a series of short consensus repeats (blue) that form a stalklike structure linked by an epidermal growth factor–like domain (yellow) to a carbohydrate-binding C-type lectin domain (red). CD62L contains a membrane-proximal site that is cleaved by a protease after neutrophil activation, resulting in shedding of the extracellular domain. (From Skubitz A, with permission.) |
Sequence of Neutrophil–Endothelial Cell Adhesion
In the Springer model, selectins are responsible for the initial rolling of the neutrophil along the endothelial cell. The association and disassociation constants of selectins for their ligands are very high, and stimulation of neutrophils can result in a rapid increase in L-selectin affinity for its ligand, resulting in tethering of a flowing cell and rolling within a millisecond (479,480). This increase in affinity is transient, and, in fact, by 5 minutes after stimulation, much of the CD62L is shed from the neutrophil surface. The close interaction of the neutrophil with the endothelial cell surface, mediated by the selectins, allows the neutrophils to sense chemoattractants released from or displayed on the endothelial surface. These chemoattractants bind to specific receptors on the neutrophil surface, many of which span the membrane seven times, are coupled with G proteins, and result in transduction of signals that activate integrin-adhesive activity (Fig. 9.22). Some of the known chemoattractants for neutrophils are listed in Table 9.9. Many tissue-derived chemotactic factors form a protein family termed chemokines. These proteins have four conserved cysteines that form two disulfide bonds. The chemokine family is composed of CXC and CC chemokines. The CXC chemokines have their first two cysteines separated by a single amino acid and stimulate neutrophils, whereas the CC chemokines do not. Among the classically described chemoattractants are the n-formyl peptides. These chemoattractants were initially identified by studies of the observation that supernatants of bacterial cultures were chemotactic for neutrophils. Subsequent studies identified a number of n-formyl peptides with chemotactic activity, and it was hypothesized that the presence of such a receptor would provide a preimmune receptor for the neutrophil to sense bacterial infections, because bacterial protein synthesis begins with n-formyl methionine, whereas mammalian protein synthesis does not. Interestingly, mammalian mitochondria do synthesize n-formyl methionyl peptides, and these may in some cases also result in neutrophil activation. Another classic chemoattractant is C5a, a cleavage fragment of the fifth component of complement.
|
|
|
Figure 9.21. Schematic of an α,β2 integrin dimer, showing the inactive, low-affinity, state (left) and the active, high-affinity, state (right). Integrins are heterodimers of and α and β subunits with a globular “head” region and two “legs.” The head of the α subunit contains EF hand repeats that are divalent metal-binding sites, and a “β propeller” domain with an I domain that contains a binding site for Mg2+ and Mn2+. The β subunit contains an I-like domain. In the low-affinity, inactive, state with no ligand bound, the integrin is bent toward the membrane with the head domain facing toward the membrane. With activation, the integrin straightens and rotates the head region outward with an associated change to higher ligand affinity. This change in structure is associated with a separation of the α and β subunits. (From Skubitz A, with permission.) |
|
|
|
Figure 9.22. Neutrophil adhesion to inflamed endothelium follows three sequential steps. First, selectin molecules (green) that recognize carbohydrate ligands bind their ligand and result in tethering and rolling along the vascular wall, bringing the neutrophil in close proximity to the endothelial cell surface. Chemotactic agents (red balls) released from the site of inflammation (red) and bound to or released from the endothelial cell surface interact with specific receptors that span the neutrophil membrane seven times and transduce activation signals via G proteins (purple) that activate integrins (black). These integrins then bind their ligands (immunoglobulin superfamily members, orange) on the endothelial cell surface, resulting in arrest of the neutrophil and subsequent transmigration across the endothelial surface and subsequent chemotaxis to the site of chemoattractant production. (From Skubitz L, with permission.) |
|
Table 9.9 Neutrophil Chemoattractants |
||
|
Integrins form a family of adhesive molecules whose affinity for ligand can be rapidly regulated. Stimulation of neutrophils with n-formyl peptides or C5a results in rapid up-regulation of CD11b/CD18 expression on the neutrophil surface. This increase in expression is caused by fusion of secretory vesicles with the cell membrane. With strong stimulation, it is possible that secondary/tertiary granule fusion contributes as well. However, mere translocation to the cell surface with an increase in cell-surface expression of CD11/CD18 is not sufficient for an increase in adhesiveness. Similarly, studies of cytoplasts have demonstrated that alterations in β2 integrin–mediated cell adhesion can be manifest without a change in surface expression of CD11b/CD18. Data suggest that alterations in CD11b/CD18-mediated cell adhesiveness are the result of a conformational change of the integrin, causing an alteration in ligand binding (148,481). Studies show that, after activation of neutrophils by chemoattractants, approximately 10% of the surface CD11b/CD18 molecules express an activation epitope recognized by a monoclonal antibody (148).
As shown in Table 9.8, several Ig superfamily members are expressed on endothelial cells and are ligands for leukocyte integrins. Mac-1 (CD11b/CD18) binds to a specific site in the third Ig domain of ICAM-1. Leukocyte function antigen-1 (CD11a/CD18) binds to the N-terminal domains of both ICAM-1 and ICAM-2. Thus, the model for neutrophil adhesion and transmigration through vessel walls can be depicted as in Figures 9.22 and 9.23. The initial rolling of neutrophils along the vessel wall is mediated by selectins (L-selectin, E-selectin, and P-selectin), and their expression and affinity for ligand can be regulated by inflammatory stimuli. At sites of inflammation, leukocyte rolling along the vessel wall is increased, and cells may become more closely apposed to the vessel wall, allowing better interaction with chemoattractants released from or presented on the surface of the endothelial cells. Interactions of these chemoattractants with the neutrophil then result in activation of integrin affinity for its ligand, with a resultant firm adhesion of the neutrophil to the endothelial cell surface. Subsequent migration of the neutrophil through the endothelial cell proceeds along the gradient of chemotactic agent. Extravasation via transcellular (through the endothelial cells) has been demonstrated, and the endothelial cell may play a role in this process (482). The relative contribution of transcellular and intercellular extravasation is unclear. The presence of multiple adhesion molecules and ligands on both the neutrophil and the endothelial cell, which may vary among endothelial cells in different environments, coupled with the array of chemoattractant agents that may be released locally, provides potentially high specificity for localizing the interaction of a particular type of cell within a particular endothelial environment, based on the large number of combinatorial adhesive molecule–ligand pair combinations available (479).
This model is supported by elegant studies demonstrating that at physiologic shear stress, neutrophils form rolling adhesions on phospholipid bilayers containing P-selectin but not on those containing ICAM-1. Chemoattractants result in integrin-mediated adhesion to bilayers containing ICAM-1 under static conditions but not under shear conditions. In contrast, neutrophils rolling on bilayers containing both P-selectin and ICAM-1 respond to chemoattractants by spreading and becoming firmly adherent via an integrin–ICAM-1 interaction. Chemoattractants do not increase adhesion or rolling on bilayers containing P-selectin alone (479).
|
|
|
Figure 9.23. Neutrophil adhesion to the vascular wall at a site of inflammation. Sequential interactions of neutrophils with selectins results in rolling along the vascular wall, followed by sensing chemoattracts (red balls), which activates integrins to cause increased adhesion to the endothelial cells, followed by transmigration through the endothelial cells and basememt membrane, followed by chemotaxis to the site of inflammation. (From Skubitz L, with permission.) |
Neutrophil Aggregation
The increase in polymorphonuclear neutrophil adhesion after stimulation is manifest not only by increased adhesion to endothelial cells, but also by neutrophil–neutrophil and neutrophil–platelet adhesion. Although the in vivo formation of neutrophil aggregates was clearly visualized by W. B. Wood in a rabbit ear model of inflammation in the 1960s, the possibility of neutrophil homotypic aggregation was considered novel when it was formally demonstrated by Craddock et al. (265,266). Craddock’s observations stemmed from earlier descriptions of the phenomenon of hemodialysis neutropenia (263,264). The initiation of hemodialysis, using cuprophane membranes, is followed by a rapid fall in the circulating neutrophil count caused by a transient sequestration of neutrophils in the lung, with a return to the circulation by 1 hour (263,264). Craddock demonstrated that neutrophils undergo homotypic aggregation in response to plasma that had been exposed to cuprophane, largely because of generation of C5a by complement activation (265,266,268,269). Aggregation was also induced by other neutrophil stimuli. Further studies demonstrated that the transient nature of hemodialysis neutropenia was caused by desensitization of neutrophils to the continued infusion of stimulus from the hemodialysis machine, thus demonstrating in vivo the phenomenon of desensitization (267). This phenomenon, in some clinical situations (e.g., viral infections), may result in neutrophil dysfunction as described later in this book. Subsequent studies have suggested that neutrophil aggregation and sludging, with resultant organ damage or dysfunction, may play a role in a variety of pathologic processes including adult respiratory distress syndrome and reperfusion injury.
Chemotaxis
The work of von Recklinghausen and Conheim described amoebalike movement of leukocytes more than a century ago (297). The neutrophil moves on a surface through a gradient of chemotactic agent by advancing a projection called a lamellipodium or pseudopodium. Chemotaxis begins with the protrusion of a pseudopodium or lamellipodium at the front of the cell. This occurs where the submembranous actin filament network (the cortex) becomes less filamentous. As the cell moves, the pseudopodium ruffles rapidly. Part of the pseudopodium adheres to the underlying surface, and the contents of the cell move forward into the pseudopodium, making the pseudopodium less prominent. This cycle is then repeated with the protrusion of another pseudopodium. Chemotaxis occurs by repetitions of this process, although often the process is so well coordinated as to appear as a continuous gliding motion. The mechanism of these cell movements has been reviewed (49) and appears to involve alterations in the polymerization state of actin, regulated by several proteins, including actin-binding protein, gelsolin, and others, as well as adenosine triphosphate–dependent contraction of the actin network mediated by myosin. Local contraction of the cytoskeleton could move intracellular components forward into an area where the cortical gel has weakened because of shortening of actin filaments beneath the surface of the advancing pseudopodium. Characteristic contraction waves have been observed in human leukocytes and likened to those seen in amebae and earthworms (Fig. 9.24) (483). In leukocytes, the contraction wave appears to originate in the superficial layer of the submembranous organelle-excluding region called the cortex, producing a concave area, and the anterior part of the cell stretches or is propelled forward as a pseudopodium (483).
|
|
|
Figure 9.24. Scanning electron micrograph of a moving neutrophil. The contraction wave is observed as a concave (black solid arrow) and a convex (black dashed arrow) area. The advancing pseudopodium (PP) is seen being pushed out in the direction of movement (white arrow). Pseudoflagellae (PF) are seen in the rear of the cell. (From Senda N, et al. The mechanism of the movement of leucocytes. Exp Cell Res 1975;91:393, with permission.) |
Interestingly, mice that express no gelsolin can breed in captivity and have a prolonged bleeding time and abnormal neutrophil chemotaxis (49). Thus, gelsolin is important in neutrophil chemotaxis, but other proteins can compensate to some extent in its absence. The increase in free calcium that alters the cytoskeleton by activating gelsolin, and thereby decreasing filamentous actin with a resultant decrease in viscosity, may play a role in locomotion; in addition, the transient dissolution of the submembranous cytoskeletal network may allow closer contact of intracellular granules with the plasma membrane, facilitating granule fusion and release. Some granule release occurs with chemotaxis.
Although the mobility of neutrophils and their concentration in inflammatory lesions were appreciated in early experiments, the development of a two-compartment chamber separated by a leukocyte-permeable membrane has permitted quantitation of chemotaxis in vitro and facilitated the investigation of chemotactic factors (484,485). Such studies revealed that neutrophils show directional migration under the influence of chemotactic agents, but a concentration gradient is needed for migration to occur. Even in the absence of a gradient, however, in the presence of a chemotactic factor, random migration is enhanced, and localization, or trapping, of the phagocytes occurs.
Phagocytosis
Metchnikoff played an important role in describing the phenomenon of phagocytosis (Fig. 9.25). When a neutrophil meets a particle, it envelops it with pseudopodia, which fuse around it, forming a phagosome that rapidly fuses with azurophilic and specific granules. Endocytosis is the process by which material is taken into a cell enclosed within pieces of plasma membrane and, therefore, never occurring free within the cytoplasm of the cell (486). Endocytosis is further divided into pinocytosis (drinking by cells) and phagocytosis (eating by cells). Phagocytosis is usually visible by light microscopy, whereas pinocytosis is not, involving ingestion of small particles, such as macromolecules. Both processes proceed through invagination of the cell membrane and the formation of vesicles or vacuoles (phagosomes).
Neutrophils and macrophages are motile and thus are free to migrate into sites of inflammation. Once they are in the area of inflammation, they come in contact with the foreign material, engulf it, and subject it to the microbicidal and digestive enzymes they contain. This sequence was appreciated by Metchnikoff in the 1880s (487), when he observed the migration of phagocytes into areas of tissue damage in sponges and lower animals. How phagocytes distinguish foreign particles and damaged autologous cells from normal self-components remains unclear, but this capacity is critical to effective phagocytic function.
|
|
|
Figure 9.25. Diagram of endocytosis; both phagocytosis of immunoglobulin-coated bacteria and pinocytosis are shown. The fusion of a primary lysosome and a specific granule with the phagosome to form the digestive vacuole, the subsequent degradation of the bacteria leading to the formation of a residual body, and the expulsion of indigestible components are also depicted. |
Granule Release
Neutrophils contain four well-defined types of intracellular granules: azurophilic, specific, and gelatinase granules, and secretory vesicles. The azurophilic granules contain many antibacterial compounds, and it appears that the fusion of these granules with phagocytic vesicles is important in bacterial killing. Azurophilic granules also contain compounds, such as elastase, that may alter locomotion by hydrolyzing certain extracellular matrix components. The specific granules are more readily released from the cell, suggesting an important function in the extracellular milieu. For example, specific granules contain products that activate the complement cascade (488). Collagenase may be important in hydrolyzing extracellular matrix proteins and facilitating locomotion. Apolactoferrin, which binds iron, may exert an antibacterial effect by depriving bacteria of iron, altering hydroxyl radical formation, and altering cell adhesion (39,40). The tertiary granules contain gelatinase in addition to other components, and, like collagenase, this enzyme may play a role in extracellular matrix remodeling during locomotion. Finally, both tertiary granules and the secretory vesicles contain membrane proteins that can be rapidly up-regulated to the cell surface and may play a role in alterations of the functional use of these surface proteins after stimulation. Membrane components of secondary granules are also up-regulated during granule release and may play a role in regulating the expression of these membrane proteins on the cell surface. The specific granules are more readily released than the azurophilic granules, and their secretion, therefore, must be regulated somewhat differently.
Bacterial Killing and Digestion
Bacterial killing by neutrophils can be ascribed to two general and often synergistic mechanisms: oxidative and nonoxidative. Bacterial killing in the phagosome is augmented by the generation of superoxide. Activated neutrophils produce superoxide via a multicomponent nicotinamide adenine dinucleotide phosphate (NADPH)–dependent oxidase that is activated by neutrophil stimulation. In resting cells, the oxidase components are found in both the plasma membrane and intracellular stores. After stimulation, intracellular components are translocated to the plasma membrane and activated, producing O2. Subsequent reactions result in the formation of H2O2 and hypochlorous acid (HOCl), which increase bacterial killing. Small amounts of other species (such as singlet oxygen and hydroxyl radical) may also form but are probably of little import in bacterial killing.
Bacterial killing decreases under anaerobic conditions, whereas phagocytosis does not, so the respiratory burst is important to bactericidal activity. Furthermore, because chronic granulomatous disease is one of the most severe clinical disorders characterized by a defect in bacterial killing, and the defect in this disorder is an inability to develop all of the reactions associated with the respiratory burst, the oxygen-dependent mechanisms appear to be of major importance in bacterial killing (489,490). However, other bactericidal mechanisms that do not require oxygen also operate within phagocytes (Table 9.9).
Oxygen-Dependent Antimicrobial Systems
Neutrophil activation is accompanied by a prominent increase in O2 use called the oxidative burst or respiratory burst, described by Baldridge and Gerard in 1933 (491). In 1964, Rossi and Zatti suggested that the respiratory burst is due to an NADPH oxidase (492); and in 1973, Babior et al. reported the production of superoxide by the NADPH oxidase (493). The respiratory burst or oxidative burst is a series of metabolic events that takes place when phagocytes are appropriately stimulated, resulting in an increase in oxygen consumption, the production of superoxide (O2-), the production of H2O2, and an increase in glucose oxidation via the hexose monophosphate shunt (489,494,495). Most of the oxidative burst is caused by activation of an NADPH oxidase that catalyzes the one-electron reduction of oxygen to superoxide, using the electron donor NADPH (489,496):
2O2 + NADPH → 2O2- + NADP+ + H+
Activation of the hexose monophosphate shunt occurs because of the increased NADP+ produced.
The NADPH oxidase exists in a latent state consisting of both membrane and cytosolic components in distinct subcellular compartments. Activation involves multiple steps, including assembly at the membrane of two membrane-bound components (gp91phox and p22phox), three cytosolic components (p40phox, p47phox, and p67phox) (the term phox indicates that the protein is a component of the phagocyte oxidase), and a low-molecular-weight G protein (rac1 or rac2) to form the membrane-bound oxidase complex (495,497,498) (Figs. 9.26 and 9.27). The activated oxidase is readily detected by nitroblue tetrazolium or cytochrome reduction or the production of chemiluminescence; several mechanisms by which this series of oxygen-dependent reactions may kill bacteria have been postulated (Table 9.10). When stimulated by phagocytosis, reactive oxygen metabolites are found localized at the phagosome, and not on parts of the plasma membrane that are not involved in phagocytosis (498). Further details of the NADPH oxidase can be found in Chapter 13.
|
|
|
Figure 9.26. Model of flavocytochrome b558. Both gp91phox and p22phox are shown, with the aligned bis-hemes and the sites for FAD and NADPH binding on gp91phox indicated. The seven specific mutations identified in patients with chronic granulomatous disease and associated with defective oxidase assembly are indicated in red bursts. Regions in the cytoplasmic loops of gp91phox that have been implicated in mediating oxidase assembly by studies using synthetic peptides or specific monoclonal antibodies are indicated in partially filled rectangles. (From Histochem Cell Biol 2004;122: 277–291, with permission.) |
Although O2- has some antibacterial activity, most O2- is rapidly converted to H2O2 by dismutation, either spontaneously or catalytically by superoxide dismutase:
2O2- + 2H+ → O2 + H2O2
Of the microbicidal oxidants generated by the respiratory burst, O2- and H2O2 are not potent microbicides; rather, they function as starting materials to generate more potent oxidizing radicals, such as oxidized halogens and oxidizing radicals (489).
Myeloperoxidase-Mediated Oxygen-Dependent Bacterial Killing with Oxidized Halogens
The antibacterial effect of hypochlorites was demonstrated by Koch in 1881, and chlorine-based disinfectants have been widely used since, though their use was suggested even earlier in the 19th century by both Alcock and Semmelweis (499). MPO is present in high concentration in the azurophilic or primary granules of neutrophils and is released into the phagosome during granule– phagosome fusion. MPO, together with H2O2 generated during phagocytosis (500,501) and an oxidizable cofactor such as halide (e.g., Cl- or Br-), forms oxidized halogens that are potent antimicrobials effective against bacteria, fungi, viruses, mycoplasma, and tumor cells (490):
Cl- + H2O2 → H2O + OCl-
The combination of MPO, halide, and H2O2 is efficient in killing bacteria at H2O2 concentrations as low as 10 μm, whereas H2O2 in the absence of MPO requires 0.5 mmol/L or greater levels to produce similar killing (489). Thus, H2O2 alone is a weak antimicrobial. Several mechanisms that have been proposed to explain bacterial killing by this system include halogenation of the bacterial cell wall, oxidation of various bacterial components, the decarboxylation of bacterial wall amino acids (489,490), and the generation of long-lived chloramines that have antimicrobial activity (489,502):
OCl- + RNH3 → OH- + RNH2Cl
|
|
|
|
Figure 9.27. Oxidase assembly. Two models for the organization of the cytosolic oxidase complex in resting phagocytes are shown. A: In the first model, p40phox provides the organizing central structure, interacting via its PC motif with the PB1 region of p67phox, and via its SH3 domain with the PRR of p47phox. B: In the second model, p67phox is at the center of the complex, bound to p40phox in the same fashion as in A but associated via its C-terminal SH3 domain with the PRR of p47phox. C: In the activated phagocyte, the cytosolic components associate at the phagosomal or plasma membrane via several contact points (bidirectional arrows). With stimulation of the cell, RacGDP dissociates from RhoGDI, undergoes guanine nucleotide exchange, and the RacGTP associates with the membrane via its C-terminal prenyl group and associates directly with gp91phox (1). p47phox, released from its autoinhibited conformation by multiple phosphorylations in the polybasic region, associates via its N-terminal SH3 domain with the PRR of p22phox (2) and via its PX domain with the phophoinositides in the target membrane (3). The TPRs of p67phox associate with RacGTP (4) while its activation domain binds directly to pg91phox (5) and its PRR associates with the C-terminal SH3 domain of p47phox (6). (From Histochem Cell Biol 2004;122: 277–291, with permission.) |
|
|
Table 9.10 Antimicrobial Systems of the Neutrophil |
|
|
There is some evidence that reactive nitrogen intermediates may also be produced (503,504).
Regardless of the mechanisms of killing, the fact that azide inhibition of MPO greatly decreases the microbicidal activity of normal leukocytes provides strong evidence for the importance of this system (500). In patients with MPO deficiency (which is often quantitative rather than qualitative), the activity of other antimicrobial systems is increased, thus partially compensating for the MPO deficiency (500). This finding may explain the increased susceptibility to infection in only approximately 20% of MPO-deficient patients (505).
Myeloperoxidase-Independent (but Oxygen Radical–Dependent) Bacterial Killing
This antimicrobial system is important because cells with no detectable MPO activity retain antibacterial actions that require oxygen; in fact, bacterial killing in MPO-deficient cells is associated with greater oxygen consumption than in normal cells (490).
Hydrogen Peroxide
As mentioned, H2O2 at high concentrations (>0.5 mmol/L) has antimicrobial activity in the absence of MPO (506). Some organisms are more sensitive than others to H2O2, and this sensitivity may depend in part on their ability to degrade it (i.e., catalase or peroxidase content). Certain substances such as iodide or ascorbic acid may enhance the bactericidal action of H2O2 or render organisms more sensitive to still other killing mechanisms, such as lysozyme (490).
Superoxide Anion
After the discovery that O2- was generated in phagocytes, some authors postulated that O2- itself might be microbicidal. The microbicidal activity of O2- appears to be weak, however, when compared to that of the H2O2 formed from it, especially if MPO is present. Superoxide by itself has minimal bactericidal activity (489,490).
Hydroxyl Radicals
Human neutrophils and monocytes generate OH· radicals, but the role of this highly reactive compound in microbial killing is unclear (489,490). OH· production from H2O2 and superoxide is catalyzed by iron in the Haber-Weiss reaction:
H2O2 + O2- → OH· + OH- + O2
OH· could also be produced from a reaction between ozone and H2O2 (497).
Singlet Oxygen
Singlet oxygen (1O2) is a highly reactive form of oxygen that is capable of attacking compounds containing double bonds. However, significant amounts of singlet oxygen are unlikely to form, so its role in bacterial killing is probably not significant (505).
Nitric Oxide
Nitric oxide may interact with neutrophil-derived oxidants to yield other relevant oxidant species, though its role in neutrophil function is unclear (499). Nitric oxide can react with superoxide to produce reactive nitrogen intermediates that can react with a variety of biologic targets. Though the clinical significance of neutrophil-derived reactive nitrogen intermediates is unclear, MPO potentiates NO-mediated nitrosation (507). Nitric oxide (NO·) reacts with superoxide (O2-·) to form the potent oxidant peroxynitrate (ONOO-). Nitrite (NO2-), a major end product of nitric oxide metabolism, has been found to interact with hypochlorous acid (HOCl) or MPO, or both, to form nitrylchloride (NO2Cl) and thus promotes tyrosine nitration (508,509). Activated neutrophils can convert NO2- to NO2Cl and NO2· through an MPO-dependent pathway and inactivate endothelial angiotensin-converting enzyme (508). Thus, neutrophil conversion of NO2- to nitrating and chlorinating species may play important physiologic roles. Nitric oxide synthase has been found in primary granules of resting neutrophils (510), and NO· production by neutrophils has been observed (509). The expression of the NOS2 isoform can be induced in neutrophils by endotoxin, whereas NOS1 is present in the resting state (511,512).
Oxygen-Independent Antimicrobial Systems
Because an anaerobic environment does not abolish antimicrobial activity, other mechanisms must be operative, and several have been identified, including the effects of acid, lysozyme, lactoferrin, defensins, cationic proteins, and neutral proteases. The delivery of the wide array of antibacterial compounds to the phagosome by fusion with azurophilic and specific granules generally results in bacterial killing caused by the direct actions of the granule contents. In addition, these effects are potentiated by the acidification of the phagosome, caused partly by the granule contents themselves, as well as by active translocation of H+ ions into the phagosome by ion pumps. The effectiveness of these mechanisms in the absence of superoxide production is demonstrated by both bacterial killing in anaerobic environments and killing by cells from patients with chronic granulomatous disease, in which catalase-positive organisms have an advantage over catalase-negative species. Nonoxidative killing is of obvious importance in hypoxic environments such as an abscess.
Acid
After particle ingestion, the intraphagosomal pH has been reported to decrease to between 3.0 and 6.5 (513,514,515). Some organisms, such as pneumococci, are sensitive to an acid pH, whereas others tolerate acid environments without damage. In addition, the acid environment may enhance the effect of lysosomal hydrolytic enzymes, most of which have optimal activity at acid pH.
Lysozyme
Lysozyme, a low-molecular-weight (14,500-dalton) basic protein, is present in both primary and secondary neutrophil granules and is capable of hydrolyzing the cell wall of certain bacteria. Most organisms are resistant to the direct action of lysozyme (490), although they may become sensitive to its action after exposure to antibody and complement or to H2O2 and ascorbic acid (490). Usually, bacterial death appears to precede the action of lysozyme, so its action may be mostly digestive. The leukocytes of Guernsey and Hereford cattle contain no lysozyme but kill organisms normally (516).
Lactoferrin
Lactoferrin is a microbiostatic protein (molecular weight 77,000 daltons) that is found in the specific granules of rabbit heterophils (517) and in human neutrophils (518) as well as in many secretions (e.g., milk and mucus) and exudates (519). It inhibits bacterial growth by binding the essential nutrient iron (two atoms per molecule), and, in contrast to transferrin, this property is maintained at the low pH values encountered in exudates. A synergistic relationship between lactoferrin and other antimicrobial systems may exist, and lactoferrin may be bactericidal for some organisms (520).
Defensins
Prominent among the cationic neutrophil granule proteins are the defensins. These small microbicidal peptides kill a variety of bacteria, fungi, and viruses (19,20,21,521). Defensins appear to exert their effects by forming voltage-dependent ion channels. They are present in very high concentration compared to other stored antibacterial peptides (approx. 5% of total neutrophil weight).
Bactericidal Permeability-Increasing Protein
Bactericidal permeability-increasing protein has antibacterial activity against certain gram-negative bacteria (24,25,26,27,28). It also has the property of neutralizing the toxic effects of endotoxin.
Other Granule Proteins
Leukocyte granules from humans, rabbits, guinea pigs, and chickens contain several other basic proteins that migrate toward the cathode on electrophoresis in agarose and exhibit antimicrobial activity (24,518). These proteins differ from species to species (24,518), and their relative importance as antimicrobial agents probably also varies. For example, because chicken leukocytes lack MPO, the cationic proteins presumably are of greatest importance in that species. In rabbit heterophils (522) and in chicken (523) and human (19,20,21) polymorphs, the cationic proteins are located in the primary granules and are delivered into the phagosome, where they coat the bacteria and are presumed to kill them (24,518). Other antibacterial granule components include azurocidin (29,30) and the serine proteinases elastase, cathepsin G, and proteinase 3 (31,32,33,34,524).
Digestion
Digestion of bacteria is demonstrated both by changes in the morphologic appearance of organisms after phagocytosis and by the release of labeled fragments of bacteria into the surrounding medium (524,525). Digestion is thought to result from the action of the acid hydrolytic enzymes released into the phagosome from the primary lysosome. Metabolic blocking agents, such as iodoacetate, cyanide, and arsenite, which inhibit glycolysis and respiration, have no effect on digestion once the bacteria are within the cell (526). Some bacteria ingested by neutrophils (e.g., certain pneumococci) may be killed and digested slowly, the undigested material remaining as myelin or residual bodies.
Unsuccessful Ingestion, Killing, or Digestion
Phagocytosis and bacterial killing are not always completed successfully. Some organisms (e.g., certain virulent staphylococci) may survive and multiply within neutrophils and appear to kill them, thus overcoming the defense mechanism (527). Still other materials ingested by neutrophils, such as the uric acid crystals of gout or the hydroxyapatite crystals of pseudogout, may cause breakdown of the phagosome wall and release the hydrolytic enzymes into the cell sap (528). This action may be fatal to the cell, which then lyses and releases its enzymes into the surrounding tissues, where they cause tissue damage and secondary inflammation. In certain streptococcal and other infections, bacterial exotoxins (e.g., streptolysin) are released and damage the phagosomal membrane, thus killing the cell in a similar manner (529); the infecting organism is freed in the process. Also, certain vitamins (vitamin A) and drugs, when incorporated into phagosomal membranes, render the membranes fragile and readily susceptible to rupture, thereby leading to inflammation (528).
Secretory Functions of the Neutrophil
In addition to the fact that the contents of the neutrophil are released passively as a result of cell lysis, a variety of substances probably are actively secreted by leukocytes in vitro. Most of these substances have been shown to originate from the granule (including secretory vesicle) fraction. Specific granule contents (lactoferrin, B12-binding protein, or both) are released before primary granule contents, and tertiary granules and secretory vesicles are secreted even more rapidly and completely, providing evidence for a differential secretion of granule contents (530). Because some of these substances are present in plasma normally and the concentration increases in patients with diseases involving the neutrophil system (531,532,533), some authors suggest that neutrophils may serve a secretory function as well as a phagocytic role in vivo (534,535).
Two modes of enzyme release or exocytosis are (a) released into phagocytic vacuoles (including release outside the cell during phagocytosis but before the phagosome is sealed off from the exterior of the cell or release during attempted phagocytosis that cannot be completed because of particle size) (530), and (b) granule content release also occurs that is not associated with phagocytosis—that is, true secretion (530).
Two well-studied released granule proteins are the B12-binding proteins or transcobalamins. Granulocytes contain and actively release B12-binding protein (534,536). This protein was thought to be a storage protein and is a poor source of metabolically available vitamin (537). It appears that transcobalamin III is derived from granulocytes; it is unsaturated with B12 (538). Markedly elevated transcobalamin I levels are seen in cases of chronic myelocytic leukemia and myeloid metaplasia; low values occur in patients with chronic leukopenia and aplastic anemia (537), and good correlation with blood granulocyte pool size has been reported (531).
Lysozyme is present in primary, secondary, and tertiary granules and is also present in monocytes, serum, and tears and other secretions (533,539). Increased concentrations in serum and urine are found in association with monocytic and myeloblastic leukemias (532,539). Although it was proposed that serum lysozyme may provide a measure of GTR (540), lysozyme is present in several cell types, and the GTR does not correlate with serum lysozyme levels in neutropenic patients. In addition, the plasma kinetics of lysozyme do not mirror the kinetics of other neutrophil granule proteins or short-term alterations in the number of circulating neutrophils (in contrast to the kinetics of lactoferrin and gelatinase) (533).
Stimulated neutrophils also synthesize and release a variety of cytokines that may regulate the inflammatory response. For example, neutrophils stimulated with lipopolysaccharides synthesize IL-1, TNF-α, and IL-1 receptor antagonist (541), whereas GM-CSF induces synthesis of TNF-α and IL-6 (542,543).
Some granule proteins originally viewed as primarily antimicrobial agents may have other effects as well. For example, neutrophil serine proteases also appear to play a regulatory role in granulopoiesis by antagonizing growth-factor effects (544,545,546). The defensins (HNP-1 to -3) also can regulate lipoprotein metabolism by stimulating the binding of lipoprotein (a) and low-density lipoprotein to bascular cells, and can regulate smooth muscle cell contraction (547).
Other Effects of Oxygen Metabolites
Reactive oxygen metabolites may also contribute to the physiologic effects of activated neutrophils by activating latent enzyme activities of granule proteases such as collagenase (548). The significance of this process for bacterial killing is unclear but is relevant to other pathophysiologic processes.
Superoxide produced by neutrophils may also stimulate fibroblast proliferation and wound healing, lymphocyte proliferation (549,550), and also may regulate gene expression and the function of some enzymes including protein kinases (reviewed in 551).
Neutrophil Antigens
Neutrophil antigens have been identified by the use of both monoclonal antibodies and patient sera using classic blood-banking techniques. Some neutrophil antigens defined by monoclonal antibodies are shown in Table 9.2. Neutrophil antigens relevant to blood banking and immune neutropenia are discussed more fully in Chapter 61. The first clinically relevant neutrophil antigens were described by Lalezari et al. and termed NA1 and its allele NA2, and a second antigen termed NB1. The NA1 and NA2 alleles were found to be present on the glycosyl phosphatidylinositol-linked receptor FcγRIIIB (552,553). The NB1 antigen (HNA-2a) (554) is present on an N-glycosylated, glycosyl phosphatidylinositol–linked 58- to 64-kDa protein of unknown function that is present in secondary granules as well as the cell surface (555,556,557).
Subsequently, a new nomenclature was established (558,559). In this system the antigens are termed HNA, for human neutrophil antigen, with the protein/antigen denoted by an integer, and the epitope by a letter. In this system NA1 became HNA-1a, NA2 became HNA-1b, and NB1 became HNA-2a (Table 9.11). Most clinically relevant allo- and autoantibodies appear to react with the HNA-1 and HNA-2 systems. Some differences in neutrophil function have been reported based on HNA phenotype. For example, neutrophils that are homozygous for HNA-1b have a lower affinity for IgG3, and phagocytose targets opsonized with IgG1 and IgG3 at a lower rate than neutrophils homozygous for HNA-1a (560,561,562). HNA-1a is typically expressed on approx. 45 to 65% of circulatory neutrophils (563,564) but on approx. 90% of circulating neutrophils in healthy people receiving G-CSF for several days (565).
Infections That Exhibit Tropism for Neutrophils
Granulocytic ehrlichiosis is a human pathogen (566,567,568,569). The Ehrlichia are obligate intracellular bacteria related to rickettsia. Human granulocytic ehrlichiosis is an acute febrile illness accompanied by severe myalgias and headaches, usually occurring within 2 weeks of contact with ixodid ticks. Common laboratory findings include leukopenia, thrombocytopenia, and increased transaminases. Although most patients respond promptly to doxycycline, death occurs in approximately 5% of reported cases, and complications such as pneumonia, renal failure, and central nervous system damage have been reported. Characteristic intracytoplasmic inclusions in neutrophils (morulae) are not always seen or recognized. Human granulocytic ehrlichiosis is closely related to two veterinary pathogens that infect granulocytes, Ehrlichia equi and Ehrlichia phagocytophila, which affect horses and ruminants, respectively.
|
Table 9.11 Neutrophil Antigens |
||||||||||||||||||||||||||||||
|
Francisella tularensis, a gram-negative bacterium that causes tularemia, can evade intracellular killing when ingested by neutrophils, in part, by disrupting the respiratory burst (570). In neutrophils infected with live F. tularensis, NADPH oxidase assembly was disrupted and the cells did not generate reactive oxygen species. At the same time, F. tularensis also impaired neutrophil activation by heterologous stimuli. Later in infection, the bacteria can escape the phagosome, and persist in the neutrophil cytosol for at least 12 hours (570).
References
1. Borregaard N, Heiple JM, Simons ER, Clark RA. Subcellular localization of the b-cytochrome component of the human neutrophil microbicidal oxidase: translocation during activation. J Cell Biol 1983;97:52–61.
2. Kjeldsen L, Sengelov H, Lollike K, et al. Isolation and characterization of gelatinase granules from human neutrophils. Blood 1994;83:1640–1649.
3. Oren A, Taylor JMG. The subcellular localization of defensins and myeloperoxidase in human neutrophils: immunocytochemical evidence of azurophil granule heterogeneity. J Lab Clin Med 1995;125:340–347.
4. Borregaard N, Lollike K, Kjeldsen L, et al. Human neutrophil granules and secretory vesicles. Eur J Haematol 1993;51:187–198.
5. Ducker TP, Skubitz KM. Subcellular localization of CD66, CD67, and NCA in human neutrophils. J Leukoc Biol 1992;52:11–16.
6. Kjeldsen L, Bainton DF, Sengelov H, Borregaard N. Identification of neutrophil gelatinase-associated lipocalin as a novel matrix protein of specific granules in human neutrophils. Blood 1994;83:799–807.
7. Plesner T, Ploug M, Ellis V, et al. The receptor for urokinase-type plasminogen activator and urokinase is translocated from two distinct intracellular compartments to the plasma membrane on stimulation of human neutrophils. Blood 1994;83:808–815.
8. de Haas M, Vossebeld PJM, von dem Borne AEGK, Roos D. Fcγ receptors of phagocytes. J Lab Clin Med 1995;126:330–341.
9. Borregaard N, Christensen L, Bjerrum OW, et al. Identification of a highly mobilizable subset of human neutrophil intracellular vesicles that contains tetranectin and latent alkaline phosphatase. J Clin Invest 1990;85: 408–416.
10. Kuroki M, Matsuo Y, Kinugasa T, Matsuoka Y. Three different NCA species, CGM6/CD67, NCA-95, and NCA-90, are comprised in the major 90 to 100-kDa band of granulocyte NCA detectable upon SDS-polyacrylamide gel electrophoresis. Biochem Biophys Res Commun 1992;182:501–506.
11. Kjeldsen L, Bainton DF, Sengelov H, Borregaard N. Identification of neutrophil gelatinase-associated lipocalin as a novel matrix protein of specific granules in human neutrophils. Blood 1994;83:799–807.
12. Sengelov H, Kjeldsen L, Kroeze W, et al. Secretory vesicles are the intracellular reservoir of complement receptor 1 in human neutrophils. J Immunol 1994; 153:804–810.
13. Lacal P, Pulido R, Sanchez-Madrid F, Mollinedo F. Intracellular location of T200 and Mo1 glycoproteins in human neutrophils. J Biol Chem 1988;263: 9946–9951.
14. Borregaard N, Cowland JB. Granules of the human neutrophilic polymorphonuclear leukocyte. Blood 1997;89:3503–3521.
15. Canonne-Hergaux F, Calafat J, et al. Expression and subcellular localization of NRAMP1 in human neutrophil granules. Blood 2002;100(1):268–275.
16. Faurschou M, Kamp S, et al. Prodefensins are matrix proteins of specific granules in human neutrophils. J Leukoc Biol 2005;78(3):785–793.
17. Theilgaard-Monch K, Jacobsen LC, et al. Haptoglobin is synthesized during granulocyte differentiation, stored in specific granules, and released by neutrophils in response to activation. Blood 2006;108(1):353–361.
18. Schultz J, Kaminker K. Myeloperoxidase of the leukocyte of normal human blood. I. Content and localization. Arch Biochem 1962;96:465–467.
19. Ganz T, Lehrer RI. Defensins. Pharmacol Ther 1995;66:191–205.
20. Martin E, Ganz T, Lehrer RI. Defensins and other endogenous peptide antibiotics of vertebrates. J Leukoc Biol 1995;58:128–136.
21. Levy O, Ooi CE, Elsbach P, et al. Antibacterial proteins of granulocytes differ in interaction with endotoxin. Comparison of bactericidal/permeability-increasing protein, p15s, and defensins. J Immunol 1995;154:5403–5410.
22. Lehrer RI, Ganz T, Selsted ME. Phagocytic defects. I. Abnormalities outside of the respiratory burst. Oxygen-independent bactericidal systems. Mechanisms and disorders. Hematol Oncol Clin North Am 1988;2:159–169.
23. Arnheim N, Inouye M, Law L, Laudin A. Chemical studies on the enzymatic specificity of goose egg white lysozyme. J Biol Chem 1973;248:233–236.
24. Spitznagel JK. Antibiotic proteins of human neutrophils. J Clin Invest 1990;86: 1381–1386.
25. Elsbach P, Weiss J. Prospects for use of recombinant BPI in the treatment of gram-negative bacterial infections. Infect Agents Dis 1995;4:102–109.
26. Elsbach P, Weiss J, Doerfler M. The bactericidal/permeability increasing protein of neutrophils is a potent antibacterial and anti-endotoxin agent in vitro and in vivo. Prog Clin Biol Res 1994;388:41–51.
27. Shafer WM, Martin LE, Spitznagel JK. Cationic antimicrobial proteins isolated from human neutrophil granulocytes in the presence of diisopropyl flu-orophosphate. Infect Immun 1984;45:29–35.
28. Wasiluk KR, Skubitz KM, Gray BH. Comparison of granule proteins for human polymorphonuclear leukocytes which are bactericidal toward pseudomonas aeruginosa. Infect Immun 1991;59:4193–4200.
29. Campanelli D, Detmers PA, Nathan CF, Gabay JE. Azurocidin and a homologous serine protease from neutrophils. Differential antimicrobial and proteolytic properties. J Clin Invest 1990;85:904–915.
30. Gabay JE, Scott RW, Campanelli D, et al. Antibiotic proteins of human polymorphonuclear leukocytes. Proc Natl Acad Sci U S A 1989;86:5610–5614.
31. Baggiolini M, Dewald B. The neutrophil. Int Arch Allergy Appl Immunol 1985; 76:13–20.
32. Henson PM, Johnson RM Jr. Tissue injury in inflammation. J Clin Invest 1987; 79:669–674.
33. Kao RC, Wehner NG, Skubitz KM, et al. Proteinase 3: a distinct polymorphonuclear leukocyte protease which produced emphysema. J Clin Invest 1988;82: 1963–1973.
34. Tanaka T, Minematsu Y, Reilly CF, et al. Human leukocyte cathepsin G. Subsite mapping with 4-nitroanilides, chemical modification, and effect of possible cofactors. Biochemistry 1985;24:2040–2047.
35. Kuroki M, Yamanaka T, Matsuo Y, et al. Immunochemical analysis of carcinoembryonic antigen (CEA)-related antigens differentially localized in intracellular granule of human neutrophils. Immun Invest 1995;24:829–843.
36. Skubitz KM, Campbell KD, Iida J, Skubitz APN. CD63 associates with tyrosine kinase activity and CD11/CD18, and transmits an activation signal in neutrophils. J Immunol 1996;157:3617–3626.
37. Kuijpers TW, Tool ATJ, van der Schoot EC, et al. Membrane surface antigen expression on neutrophils: a reappraisal of the use of surface markers for neutrophils activation. Blood 1991;78:1105–1111.
38. Oram JD, Reiter B. Inhibition of bacteria by lactoferrin and other iron-chelating agents. Biochim Biophys Acta 1968;170:351–365.
39. Oseas R, Yang HH, Baehner RL, Boxer LA. Lactoferrin: a promoter of polymorphonuclear leukocyte adhesiveness. Blood 1981;57:939–945.
40. Boxer LA, Haak RA, Yang HH, et al. Membrane-bound lactoferrin alters the surface properties of polymorphonuclear leukocytes. J Clin Invest 1982;70: 1049–1057.
41. Halliwell B, Gutteridge JMC. Oxygen free radicals and iron in relation to biology and medicine: some problems and concepts. Arch Biochem Biophys 1986;246:501–514.
42. Aruoma OI, Halliwell B. Superoxide-dependent and ascorbate-dependent formation of hydroxyl radicals from hydrogen peroxide in the presence of iron: are lactoferrin and transferrin promoters of hydroxyl-radical generation? Biochem J 1987;241:273–278.
43. Dewald B, Bretz U, Baggiolini M. Release of gelatinase from a novel secretory compartment of human neutrophils. J Clin Invest 1982;70:518–525.
44. Kanayama N, Kajiwara Y, Goto J, et al. Inactivation of interleukin-8 by aminopeptidase N (CD13). Blood 1995;57:129–134.
45. Painter RG, Dukes R, Sullivan J, et al. Function of neutral endopeptidase on the cell membrane of human neutrophils. J Biol Chem 1988;263:9456–9461.
46. Skubitz KM, Campbell KD, Skubitz APN. CD66a, CD66b, CD66c, and CD66d each independently stimulate neutrophils. J Leukoc Biol 1996;60:106–117.
47. Harvath L, Balke JA, Christiansen NP, et al. Selected antibodies to leucocyte common antigen (CD45) inhibit human neutrophil chemotaxis. J Immunol 1991;146:949–957.
48. Southwick FS, Stossel TP. Contractile proteins in leukocyte function. Semin Hematol 1983;20:305–321.
49. Stossel TP. The machinery of blood cell movements. Blood 1994;367–379.
50. Gottfried EL. Lipids of human leukocytes: relation to cell type. J Lipid Res 1967;8:321–327.
51. Gottfried EL. Lipid patterns of leukocytes in health and disease. Semin Hematol 1972;9:241–250.
52. Bjerrum OW, Nielsen H, Borregaard N. Quantitative analysis of phospholipids and demonstration of plasmalogen in human neutrophil subcellular fractions by high-performance liquid chromatography. Scand J Clin Lab Invest 1989;49:613–622.
53. Samuelsson B, Dahlen SE, Lindgren JA, et al. Leukotrienes and lipoxins: structures, biosynthesis and biological effects. Science 1987;237:1171–1176.
54. Samuelsson B, Goldyne M, Granstrom E, et al. Prostaglandins and thromboxanes. Annu Rev Biochem 1978;47:997–1029.
55. Symington FW. CDw17: a neutrophil glycolipid antigen regulated by activation. J Immunol 1989;8:2784–2790.
56. Symington FW, Murray WA, Bearman SI, Hakomori SI. Intracellular localization of lactosylceramide, the major human neutrophil glycosphingolipid. J Biol Chem 1987;262:11356–11363.
57. Kiguchi K, Henning-Chubb CB, Huberman E. Glycosphingolipid patterns of peripheral blood lymphocytes, monocytes, and granulocytes are cell specific. J Biochem 1990;107:8–14.
58. Kniep B, Skubitz KM. Subcellular localization of glycosphingolipids in human neutrophils. J Leukoc Biol 1998;63(1):83–88.
59. Cinek T, Horejsi V. The nature of large noncovalent complexes containing glycosyl-phosphatidylinositol-anchored membrane glycoproteins and protein tyrosine kinases. J Immunol 1992;149(7):2262–2270.
60. Surviladze Z, Draberova L, et al. Functional heterogeneity of Thy-1 membrane microdomains in rat basophilic leukemia cells. Eur J Immunol 1998;28(6): 1847–1858.
61. Brown DA, Rose JK. Sorting of GPI-anchored proteins to glycolipid-enriched membrane subdomains during transport to the apical cell surface. Cell 1992;68(3):533–544.
62. Stefanova I, Horejsi V, et al. GPI-anchored cell-surface molecules complexed to protein tyrosine kinases. Science 1991;254(5034):1016–1019.
63. Skubitz KM, Campbell KD, et al. CD63 associates with CD11/CD18 in large detergent-resistant complexes after translocation to the cell surface in human neutrophils. FEBS Lett 2000;469(1):52–56.
64. Bozza PT, Payne JL, et al. Mechanisms of platelet-activating factor-induced lipid body formation: requisite roles for 5-lipoxygenase and de novo protein synthesis in the compartmentalization of neutrophil lipids. J Exp Med 1996;183(4):1515–1525.
65. Yu W, Cassara J, et al. Phosphatidylinositide 3-kinase localizes to cytoplasmic lipid bodies in human polymorphonuclear leukocytes and other myeloid-derived cells. Blood 2000;95(3):1078–1085.
66. Weller PF, Ryeom SW, Picard ST, et al. Cytoplasmic lipid bodies of neutrophils: formation induced by cis-unsaturated fatty acids and mediated by protein kinase C. J Cell Biol 1991;113:137–146.
67. Lemarchand P, Vaglio M, Mauel J, Markert M. Translocation of a small cytosolic calcium-binding protein (MRP-8) to plasma membrane correlates with human neutrophil activation. J Biol Chem 1992;267:19379–19382.
68. Odink K, Cerletti N, Bruggen J, et al. Two calcium-binding proteins in infiltrate macrophages of rheumatoid arthritis. Nature 1987;330:80–82.
69. Hogg N, Allen C, Edgeworth J. Monoclonal antibody 5.5 reacts with p8, 14, a myeloid molecule associated with some vascular endothelium. Eur J Immunol 1989;19:1053–1061.
70. Dale I, Fagerhol MK, Naesgaard I. Purification and partial characterization of a highly immunogenic human leukocyte protein, the L1 antigen. Eur J Biochem 1983;134:1–6.
71. Wilkinson MM, Busuttil A, Hayward C, et al. Expression pattern of two related cystic fibrosis-associated calcium-binding proteins in normal and abnormal tissues. J Cell Sci 1988;91:221–230.
72. Dorin JR, Novak M, Hill RE, et al. A clue to the basic defect in cystic fibrosis from cloning the CF antigen gene. Nature 1987;326:614–617.
73. Stroncek DF, Shankar RA, et al. The subcellular distribution of myeloid-related protein 8 (MRP8) and MRP14 in human neutrophils. J Transl Med 2005;3:36.
74. Giorgi R, Pagano RL, et al. Antinociceptive effect of the calcium-binding protein MRP-14 and the role played by neutrophils on the control of inflammatory pain. J Leukoc Biol 1998;64(2):214–220.
75. Perretti M, Wheller SK, et al. Modulation of cellular annexin I in human leukocytes infiltrating DTH skin reactions. J Leukoc Biol 1999;65(5):583–589.
76. Ezekowitz RAB, Orkin SH, Newburger PE. Recombinant interferon gamma augments phagocyte superoxide production and X-chronic granulomatous disease gene expression in X-linked variant chronic granulomatous disease. J Clin Invest 1987;80:1009–1016.
77. LaFleur M, Beaulieu AD, Kreis C, Poubelle P. Fibronectin gene expression in polymorphonuclear leukocytes. Accumulation of mRNA in inflammatory cells. J Biol Chem 1987;262:2111–2115.
78. Woodman RC, Curnutte JT, Babior BM. Evidence that de novo protein synthesis participates in a time-dependent augmentation of the chemotactic peptide-induced respiratory burst in neutrophils. Effects of recombinant human colony stimulating factors and dihydrocytochalasin B. J Free Radicals Biol Med 1988;5:355–361.
79. Breton-Gorius J, Reyes F. Ultrastructure of human bone marrow cell maturation. Int Rev Cytol 1976;46:251.
80. Orlic D, et al. An ultrastructural study of erythropoietin-induced red cell formation in mouse spleen. J Ultrastruct Res 1965;13:516.
81. Rifkind RA, et al. An ultrastructural study of early morphogenetic events during the establishment of fetal hepatic erythropoiesis. J Cell Biol 1969;40:343.
82. Tooze J, Davies HG. Light- and electron-microscope studies on the spleen of the newt Triturus cristatus: the fine structure of erythropoietic cells. J Cell Sci 1967;2:617.
83. Wetzel BK. The fine structure and cytochemistry of developing granulocytes, with special reference to the rabbit. In: Gordon AS, ed. Regulation of hematopoiesis, vol II. White cell and platelet production. New York: Appleton-Century-Crofts, 1970.
84. Hardin JH, Spicer SS. Ultrastructural localization of dialyzed iron-reactive mucosubstance in rabbit heterophils, basophils, and eosinophils. J Cell Biol 1971;48:368–386.
85. Erber WN, et al. Immunocytochemical labeling of haematological samples using monoclonal antibodies. Hamatol Bluttransfus 1985;29:180.
86. Dao C, et al. Eosinophil and neutrophil colony-forming cells in culture. Blood 1977;50:833.
87. Denburg JA, et al. Heterogeneity of human peripheral blood eosinophil-type colonies: evidence for a common basophil-eosinophil progenitor. Blood 1985;66:312.
88. Ogawa M, et al. Renewal and commitment to differentiation of hemopoietic stem cells (an interpretive review). Blood 1983;61:823.
89. Quesenberry P, Levitt L. Hematopoietic stem cells. N Engl J Med 1979;301:755, 819, 868.
90. Hayhoe FGJ, Quaglmo D. Haematological cytochemistry. Edinburgh: Churchill Livingstone, 1980.
91. Boll I, Kuhn A. Granulocytopoiesis in human bone marrow cultures studied by means of kinematography. Blood 1965;26:449.
92. Capone RJ, et al. Electron microscope studies on normal human myeloid elements. Blood 1964;23:300.
93. Bainton DF, et al. The development of neutrophilic PMN leukocytes in human bone marrow: origin and content of azurophil and specific granules. J Exp Med 1971;134:907.
94. Rich AR, et al. The differentiation of myeloblasts from lymphoblasts by their manner of locomotion. Bull Johns Hopkins Hosp 1939;65:291.
95. Senda N. The movement of leucocytes. J Clin Electron Microsc 1974;7:3.
96. Downey H. The myeloblast. In: Cowdry EV, ed. Special cytology, 2nd ed. New York: Paul B Hoeber, 1932.
97. Jordan HE. Comparative hematology. In: Downey H, ed. Handbook of hematology II. New York: Paul B Hoeber, 1938.
98. Gralnick HR, et al. Classification of acute leukemia. Ann Intern Med 1977; 87:740.
99. Ackerman GA. Ultrastructure and cytochemistry of the developing neutrophil. Lab Invest 1968;19:290.
100. Scott RE, Horn RG. Ultrastructural aspects of neutrophil granulocyte development in humans. Lab Invest 1970;23:202.
101. Bainton DF, Farquhar MG. Origin of granules in polymorphonuclear leukocytes: two types derived from opposite faces of the Golgi complex. J Cell Biol 1966;28:277.
102. Ackerman GA, Bellios NC. A study of the morphology of the living cells of blood and bone marrow in vital films with the phase contrast microscope. I. Normal blood and bone marrow. Blood 1955;10:3.
103. Lichtman MA. Cellular deformability during maturation of the myeloblast: possible role in marrow egress. N Engl J Med 1970;283:943.
104. Lichtman MA, Weed RI. Alteration of the cell periphery during granulocyte maturation: relationship to cell function. Blood 1972;39:301.
105. Nachman R, et al. Studies on isolated membranes of azurophil and specific granules from rabbit polymorphonuclear leukocytes. J Cell Biol 1972;54:133.
106. West BC, et al. Isolation and partial characterization of human eosinophil granules. Am J Pathol 1975;81:575.
107. Bainton DF, et al. Primary lysosomes of blood leukocytes. In: Dingle JT, Dean RT, eds. Lysosomes in biology and pathology, vol 5. New York: Elsevier, 1976:3. (Frontiers of Biology series, vol 45).
108. Ackerman GA. Histochemical differentiation during neutrophil development and maturation. Ann N Y Acad Sci 1964;113:537.
109. Baggiolini M. The neutrophil. In: Weissmann G, ed. The cell biology of inflammation. New York: Elsevier, 1980.
110. Dewald B, et al. Subcellular localization and heterogeneity of neutral proteases in neutrophilic polymorphonuclear leukocytes. J Exp Med 1975;141:709.
111. Bainton DF, Farquhar MG. Difference in enzyme content of azurophil and specific granules of polymorphonuclear leukocytes. J Cell Biol 1968;39:299.
112. Farquhar MG, et al. Cytochemical localization of acid phosphatase activity in granule fractions from rabbit polymorphonuclear leukocytes. J Cell Biol 1972;54:141.
113. Cramer E, et al. Ultrastructural localization of lactoferrin and myeloperoxidase in human neutrophils by immunogold. Blood 1985;65:423.
114. Lajtha LG, et al. Uptake of S35 sulphate by human bone marrow cells in vitro. Br J Cancer 1953;7:401.
115. Vodopick HA, et al. An evaluation of radiosulfate as a granulocyte label in the dog. J Lab Clin Med 1966;68:47.
116. Zeya HI, Spitznagel JK. Cationic protein-bearing granules of polymorphonuclear leukocytes: separation from enzyme-rich granules. Science 1969;163: 1069.
117. Kaplow LS. Cytochemistry of leukocyte alkaline phosphatase. Am J Clin Pathol 1963;39:439.
118. Wetzel BK, et al. Fine structural localization of acid and alkaline phosphatase in cells of rabbit blood and bone marrow. J Histochem Cytochem 1967;15:311.
119. Baggiolini M, et al. Resolution of granules from rabbit heterophil leukocytes into distinct populations by zonal sedimentation. J Cell Biol 1969;40:529.
120. Bretz U, Baggiolini M. Association of alkaline phosphatase of rabbit polymorphonuclear leukocytes with the membrane of the specific granules. J Cell Biol 1973;59:696.
121. Rustin GJS, et al. Studies on the subcellular localization of human neutrophil alkaline phosphatase. J Cell Sci 1979;36:401.
122. Hirschhorn R, Weissmann G. Isolation and properties of human leukocyte lysosomes in vitro. Proc Soc Exp Biol Med 1965;119:36.
123. Dewald B, et al. Release of gelatinase from a novel secretory compartment of human neutrophils. J Clin Invest 1982;70:518.
124. Watanabe I, et al. Method for electron microscopic studies of circulating human leukocytes and observations on their fine structure. J Ultrastruct Res 1967;20:366.
125. Payne DN, Ackerman GA. Ultrastructural autoradiographic study of the uptake and intracellular localization of 35S-sulfate by developing human neutrophils. Blood 1977;50:841.
126. Borregaard N, Sehested M, Nielsen BS, et al. Biosynthesis of granule proteins in normal human bone marrow cells. Gelatinase is a marker of terminal neutrophil differentiation. Blood 1995;85:812–817.
127. Wilkinson PC. The locomotion of leucocytes: definitions and descriptions. In: Chemotaxis and inflammation, 2nd ed. New York: Churchill Livingstone, 1982.
128. Ponder E, Mineola LI. The polycyte. J Lab Clin Med 1942;27:866.
129. Orfanakis NG, et al. Normal blood leukocyte concentration values. Am J Clin Pathol 1970;54:647.
130. Cartwright GE. Diagnostic laboratory hematology. New York: Grune & Stratton, 1968.
131. Edwin E. The segmentation of polymorphonuclear neutrophils. Acta Med Scand 1967;182:401.
132. Osgood EE, Ashworth CM. An atlas of hematology. San Francisco: JW Stacey, 1937.
133. Herbert V. Megaloblastic anemias: mechanisms and management. Chicago: Year Book, 1965.
134. Marsh JC, et al. Neutrophil kinetics in acute infection. J Clin Invest 1967;46: 1943.
135. Zucker-Franklin D. Electron microscopic studies of human granulocytes: structural variations related to function. Semin Hematol 1968;5:109.
136. Sin YM, Sainte-Marie G. Granulocytopoiesis in the rat thymus. I. Description of the cells of the neutrophilic and eosinophilic series. Br J Haematol 1965; 11:613.
137. Eastment CE, et al. The role of the nuclear protein matrix during development of rabbit granulocytes. Blood 1981;57:747.
138. Arneth J. Die Neutrophilen wiessen Blutkörperchen bei Infections-Krankheiten. Jena, Germany: Fischer, 1904.
139. Fliedner TM, et al. Das Studium der Proliferationsdynamik der Myelopose unter Verwendung der Einzellautoradiographie. Folia Haematol (Leipz) 1961; 6:210.
140. Lewis WH. On the locomotion of the polymorphonuclear neutrophils of the rat in autoplasma cultures. Bull Johns Hopkins Hosp 1934;55:273.
141. Sabin FR. Studies of living human blood cells. Bull Johns Hopkins Hosp 1923;34:277–288.
142. Fliedner TM, et al. Granulocytopoiesis. I. Senescence and random loss of neutrophilic granulocytes in human beings. Blood 1964;24:402.
143. McCutcheon M. Studies on the locomotion of leukocytes I. The normal rate of locomotion of human neutrophilic leukocytes in vitro. Am J Physiol 1923;66: 180–195.
144. Howard TH. Quantitation of the locomotive behavior of polymorphonuclear leukocytes in clot preparation. Blood 1982;59:946–951.
145. Harvath L, Leonard EJ. Two neutrophil populations in human blood with different chemotactic activities: separation and chemoattractant binding. Infect Immun 1982;36:443–551.
146. Seligman B, Malech HL, Melnich DA, Gallin JI. An antibody binding to human neutrophils demonstrates antigenic heterogeneity detected early in myeloid maturation which correlates with functional heterogeneity of mature neutrophils. J Immunol 1985;135:2647–2653.
147. Clement LT, Lehmeryer JE, Garland GL. Identification of neutrophil subpopulations with monoclonal antibodies. Blood 1983;61:326–332.
148. Diamond MS, Springer TA. A subpopulation of Mac-1 (CD11b/ CD18) molecules mediates neutrophil adhesion to ICAM-1 and fibrinogen. J Cell Biol 1993;120: 545–556.
149. Krause PJ, Malech HL, Kristie J, et al. Polymorphonuclear leukocyte heterogeneity in neonates and adults. Blood 1986;68:200–204.
150. Ross GD. Structure and function of membrane complement receptors. Fed Proc 1982;41:3089–3093.
151. Ross GD, Jarowsky CI, Rabellino EM, Winchester RJ. The sequential appearance of Ia-like antigens and two different complement receptors during the maturation of human neutrophils. J Exp Med 1978;147:730–744.
152. Genco RJ, van Dyke TE, Levine MJ, et al. Molecular factors influencing neutrophil defects in periodontal disease J. Dent Res 1986;64:1377–1391.
153. Berkow RL, Baehner RL. Volume-dependent human blood polymorphonuclear leukocyte heterogeneity demonstrated with counterflow centrifugal elutriation. Blood 1985;65:71.
154. Gallin JI. Human neutrophil heterogeneity exists, but is it meaningful? Blood 1984;63:977.
155. Scott CS, et al. C3b receptor-negative peripheral blood neutrophils. Scand J Haematol 1984;32:183.
156. Olsson I, et al. Purification of eosinophil peroxidase and studies of biosynthesis and processing in human marrow cells. Blood 1985;66:1143.
157. Bujak JS, Root RK. The role of peroxidase in the bactericidal activity of human blood eosinophils. Blood 1974;43:727.
158. Olsson I, et al. Arginine-rich cationic proteins of human eosinophil granules. Comparison of the constituents of eosinophilic and neutrophilic leukocytes. Lab Invest 1977;36:493.
159. Ackerman SJ, et al. Distinctive cationic proteins of the human eosinophil granule: major basic protein, eosinophil cationic protein, and eosinophil-derived neurotoxin. J Immunol 1983;131:2977.
160. Ward JC, Gitlin JB, Garry DJ, et al. Epidermolysis bullosa acquisita induced by GM-CSF: a role for eosinophils in treatment-related toxicity. Br J Haematol 1992;81:27–32.
161. DeMello DE, Liapis H, Jureidina S, et al. Cardiac localization of eosinophil-granule major basic protein in acute necrotizing myocarditis. N Engl J Med 1990;323:1542–1545.
162. Fureder W, Agis H, Sperr WR, et al. The surface membrane antigen phenotype of human blood basophils. Allergy 1994;49:861–865.
163. Zucker-Franklin D. Electron microscopic studies of human basophils. Blood 1967;29:878.
164. Hastie R. The antigen-induced degranulation of basophil leukocytes from atopic subjects, studied by phase-contrast microscopy. Clin Exp Immunol 1971;8:45.
165. Hastie R. A study of the ultrastructure of human basophil leukocytes. Lab Invest 1974;31:223.
166. Ackerman GA. Cytochemical properties of the blood basophilic granulocyte. Ann N Y Acad Sci 1963;103:376.
167. Hastie R, et al. The antigen-induced degranulation of basophil leukocytes from atopic subjects studied by electron microscopy. Lab Invest 1977;36:173.
168. Galli SJ, et al. Basophils and mast cells: morphologic insights into their biology, secretory patterns and functions. Prog Allergy 1984;34:1.
169. Bainton DF, et al. Formation and structure of specific granules in basophilic leukocytes of the guinea pig. Lab Invest 1969;21:65.
170. Kennedy WP, MacKay I. The macropolycyte in health and disease in Iraq. J Pathol 1937;44:701.
171. Oria J, Yoneda S. Variation of the form and structure of the nucleus in the various types of plurisegmented neutrophils. In: Dameshek W, Taylor FHL, eds. George R Minot symposium of hematology. New York: Grune & Stratton, 1949.
172. Weicker H. A quantitative model of granulopoiesis. Schweiz Med Wochenschr 1956;86:1459.
173. Davidson WM, Smith DR. A morphological sex difference in the polymorphonuclear neutrophil leucocytes. Br Med J 1954;2:6.
174. Hochstenbach PF, Scheres JM, Hustinx TW, Wieringa B. Demonstration of X chromatin in drumstick-like nuclear appendages of leukocytes by in situ hybridization on blood smears. Histochemistry 1986;84:383–386.
175. Maclean N, Edin MB. The drumsticks of polymorphonuclear leucocytes in sex-chromosome abnormalities. Lancet 1962;1:1154.
176. Davidson WM. Sexing the blood leucocytes in abnormalities of the sex chromosomes. Minerva Pediatr 1965;17:585.
177. Tomonaga M, et al. Leukocyte drumsticks in chronic granulocytic leukemia and related disorders. Blood 1961;18:581.
178. Davidson WM, Winn S. The relationship between the sex nodule and the sex chromosomes. In: Davidson WM, Smith DR, eds. Human chromosomal abnormalities. London: Staples Press, 1961.
179. Simmons A, et al. Normal laboratory values for differential white cell counts established by normal and automated cytochemical methods (Hemalog D-TM). J Clin Pathol 1974;27:55–58.
180. Bain BJ, England JM. Normal haematological values: sex difference in neutrophil count. Br Med J 1975;1:306.
181. Berrebi A, et al. Leukopenia in Ethiopian Jews. N Engl J Med 1987;316:549.
182. Shaper AG, Lewis P. Genetic neutropenia in people of African origin. Lancet 1971;2:1021.
183. Haden RL. Qualitative changes in neutrophilic leukocytes. Am J Clin Pathol 1935;5:354; also in Principles of hematology. Philadelphia: Lea & Febiger, 1939.
184. Athens JW. Neutrophilic granulocyte kinetics and granulocytopoiesis. In: Gordon AS, ed. Regulation of hematopoiesis. New York: Appleton-Century-Crofts, 1970.
185. Boggs DR. The kinetics of neutrophilic leukocytes in health and in disease. Semin Hematol 1967;4:359.
186. Cartwright GE, et al. The kinetics of granulopoiesis in normal man. Blood 1964;24:780.
187. Cronkite EP. Enigmas underlying the study of hemopoietic cell proliferation. Fed Proc 1964;23:649.
188. Cronkite EP, Fliedner TM. Granulocytopoiesis. N Engl J Med 1964;270: 1347–1403.
189. Cronkite EP, Vincent PC. Granulocytopoiesis. In: Stohlman F Jr, ed. Hemopoietic cellular proliferation. New York: Grune & Stratton, 1970.
190. Cronkite EP, Vincent PC. Granulocytopoiesis. Ser Haematol 1969;2:3.
191. LoBue J. Analysis of normal granulocyte production and release. In: Gordon AS, ed. Regulation of hematopoiesis, vol II. New York: Appleton-Century-Crofts, 1970.
192. Mauer AM, et al. Leukokinetic studies. II. A method for labeling granulocytes in vitro with radioactive diisopropylfluorophosphate (DFP32). J Clin Invest 1960; 39:1481.
193. Patt HM. A consideration of myeloid-erythroid balance in man. Blood 1957; 12:777.
194. Patt HM, Maloney MA. An evaluation of granulocytopoiesis. In: Lamerton LF, Fry RJM, eds. Cell proliferation. Philadelphia: FA Davis, 1963.
195. Patt HM, Maloney MA. Kinetics of neutrophil balance. In: Stohlman F Jr, ed. The kinetics of cellular proliferation. New York: Grune & Stratton, 1959.
196. Perry S, et al. Physiology of the granulocyte. JAMA 1968;203:937.
197. Warner HR, Athens JW. An analysis of granulocyte kinetics in blood and bone marrow. Ann N Y Acad Sci 1964;113:523.
198. Wilbur DW. The neutrophil system. Lawrence Radiation Laboratory Report. Springfield, VA: National Bureau of Standards, US Department of Commerce, 1966.
199. Cronkite EP, et al. The use of tritiated thymidine in the study of haemopoietic cell proliferation. In: Wolstenholme GEW, O’Connor M, eds. Ciba foundation symposium on haemopoiesis. London: J & A Churchill, 1960.
200. Maloney MA, Patt HM. Neutrophil life cycle with tritiated thymidine. Proc Soc Exp Biol Med 1958;98:801.
201. Bond VP, et al. Proliferative potentials of bone marrow and blood cells studied by in vitro uptake of 3H-thymidine. Acta Haematol (Basel) 1959;21:1.
202. Lord BI. Cellular proliferation in normal and continuously irradiated rat bone marrow studied by repeated labeling with tritiated thymidine. Br J Haematol 1965;11:130.
203. Price TH, Ochs HD, Gershoni-Baruch R, et al. In vivo neutrophil and lymphocyte functions studies in a patient with leukocyte adhesion deficiency type II. Blood 1994;84:1635–1639.
204. Jagels MA, Chambers JD, Arfors KE, Hugli TE. C5a- and tumor necrosis factor-α-induced leukocytosis occurs independently of β2 integrins and L-selectin: differential effects on neutrophil adhesion molecule expression in vivo. Blood 1995;85:2900–2909.
205. Jagels MA, Hugli TE. Neutrophil chemotactic factors promote leukocytosis, a common mechanism for cellular recruitment from bone marrow. J Immunol 1992;148:1119–1128.
206. Patt HM, Maloney MA. A model of granulocyte kinetics. Ann N Y Acad Sci 1964; 113:515.
207. Lajtha LG, et al. Kinetics of a bone-marrow stem-cell population. Ann N Y Acad Sci 1964;113:742.
208. Sin YM, Sainte-Marie G. Granulocytopoiesis in the rat thymus. II. Pattern for granulocyte formation based on cell counts in granulocytopoietic islands. Br J Haematol 1965;2:624.
209. Fliedner TM, et al. The mitotic index of human bone marrow in healthy individuals and irradiated human beings. Acta Haematol (Basel) 1959;22:65.
210. Killmann S, et al. Mitotic indices of human bone marrow cells. III. Duration of some phases of erythrocytic and granulocytic proliferation computed from mitotic indices. Blood 1964;24:267.
211. Killmann S, et al. Mitotic indices of human bone marrow cells. I. Number and cytologic distribution of mitoses. Blood 1962;19:743.
212. Lala PK, et al. A comparison of two markers of cell proliferation in bone marrow. Acta Haematol (Basel) 1964;31:1.
213. Leblond CP, Walker BF. Renewal of cell populations. Physiol Rev 1956;36:255.
214. Killmann S, et al. Mitotic indices of human bone marrow cells. II. The use of mitotic indices for estimation of time parameters of proliferation in serially connected multiplicative cellular compartments. Blood 1963;21:141.
215. Cronkite EP, et al. Anatomic and physiologic facts and hypotheses about hemopoietic proliferating systems. In: Stohlman F Jr, ed. The kinetics of cellular proliferation. New York: Grune & Stratton, 1959.
216. Dacie JV, White JC. Erythropoiesis with particular reference to its study by biopsy of human bone marrow: a review. J Clin Pathol 1949;2:1.
217. Japa J. A study of the mitotic activity of normal human bone marrow. Br J Exp Pathol 1942;23:272.
218. Mauer AM. Diurnal variation of proliferative activity in the human bone marrow. Blood 1965;26:1.
219. Golobova MT. Changes in mitotic activity in rats in relation to the time of day and night. Bull Exp Biol Med Trans 1958;46:1143.
220. Haus E, Nicolau GY, Lakatua D, Sackett-Lundeen L. Reference values for chronopharmacology. Ann Rev Chronopharmacol 1988;4:333–351.
221. Dancey JT, et al. Neutrophil kinetics in man. J Clin Invest 1976;58:705.
222. Deubelbeiss KA, et al. Marrow erythroid and neutrophil cellularity in the dog. J Clin Invest 1975;55:825.
223. Donohue DM, et al. Quantitative measurement of hematopoietic cells of the marrow. J Clin Invest 1958;37:1564.
224. Donohue DM, et al. Quantitative measurement of the erythrocytic and granulocytic cells of the marrow and blood. J Clin Invest 1958;37:1571.
225. Cronkite EP, et al. The use of tritiated thymidine in the study of DNA synthesis and cell turnover in hemopoietic tissues. Lab Invest 1959;3:263.
226. Feinendegen LE, et al. Physiological thymidine reutilization in rat bone marrow. Proc Soc Exp Biol Med 1966;122:448.
227. Feinendegen LE, et al. RNA turnover in normal rat bone marrow. Ann N Y Acad Sci 1964;113:727.
228. Friedkin M. Speculations on the significance of thymidylic acid synthesis in the regulation of cellular proliferation. In: Stohlman F Jr, ed. The kinetics of cellular proliferation. New York: Grune & Stratton, 1959.
229. Greulich RC, et al. Stimulation of mitosis in adult mice by administration of thymidine. Proc Natl Acad Sci U S A 1961;47:743.
230. Hiatt HH, Bojarski TB. Stimulation of thymidylate kinase activity in rat tissues by thymidine administration. Biochem Biophys Res Commun 1960;2:35.
231. Hughes WL, et al. Discussion of radiation effects of cellular labels. In: Stohlman F Jr, ed. The kinetics of cellular proliferation. New York: Grune & Stratton, 1959.
232. Samuels LD, et al. Tritiated thymidine toxicity in mammalian systems. Atompraxis 1964;18:144.
233. Cronkite EP. Kinetics of granulocytopoiesis. Natl Cancer Inst Monogr 30, Human Tumor Cell Kinetics, 1969.
234. Stryckmans P, et al. DNA synthesis time of erythropoietic and granulopoietic cells in human beings. Nature 1966;211:717.
235. Deubelbeiss KA, et al. Neutrophil kinetics in the dog. J Clin Invest 1975;55:833.
236. Brubaker LH, Evans WH. Separation of granulocytes, monocytes, lymphocytes, erythrocytes, and platelets from human blood and relative tagging with diisopropylfluorophosphate (DFP). J Lab Clin Med 1969;73:1036.
237. Kurth D, et al. Leukokinetic studies. V. Uptake of tritiated diisopropylfluorophosphate by leukocytes. Proc Soc Exp Biol Med 1964;1407:422.
238. Bishop CR, et al. Leukokinetic studies. XIV. Blood neutrophil kinetics in chronic, steady-state neutropenia. J Clin Invest 1971;50:1678.
239. Raab SO, et al. Granulokinetics in normal dogs. Am J Physiol 1964;206:83.
240. Kindred JE. A quantitative study of the hemopoietic organs of young adult albino rats. Am J Anat 1942;71:297.
241. Papayannopoulou T, Finch CA. On the in vivo action of erythropoietin: a quantitative analysis. J Clin Invest 1972;51:1179.
242. Hudson G. Bone marrow volume in guinea-pigs. J Anat 1958;92:150.
243. Hudson G, et al. Cell-populations in the bone marrow of the normal guinea pig. Acta Anat (Basel) 1963;52:234.
244. Suit HD. A technique for estimating bone marrow cellularity in vivo using 59Fe. J Clin Pathol 1957;10:267.
245. Harker LA. Megakaryocyte quantitation. J Clin Invest 1968;47:452.
246. Finch CA, et al. Kinetics of the formed elements of human blood. Blood 1977; 50:699.
247. Fliedner TM, et al. Granulocytopoiesis. II. Emergence and pattern of labeling of neutrophilic granulocytes in humans. Blood 1964;24:683.
248. Perry S, et al. Rates of appearance and disappearance of white blood cells in normal and in various disease states. J Lab Clin Med 1958;51:501.
249. Maloney MA, Patt HM. Granulocyte transit from bone marrow to blood. Blood 1968;31:195.
250. Bishop CR, Athens JW. Studies of granulocytopoiesis in abnormal conditions. In: Stohlman F Jr, ed. Hemopoietic cellular proliferation. New York: Grune & Stratton, 1970.
251. Boggs DR, et al. Leukokinetic studies. IX. Experimental evaluation of a model of granulopoiesis. J Clin Invest 1965;44:642.
252. Weiss L. The hematopoietic microenvironment of the bone marrow: an ultrastructural study of the stoma in normal and eosinophilic rats. Anat Rec 1976; 186:161.
253. Weiss L. Transmural cellular passage in vascular sinuses of rat bone marrow. Blood 1970;36:189.
254. Leblond PF, et al. Scanning electron microscopy of erythropoietin-stimulated bone marrow. Blood Cells 1975;1:639.
255. Giordano GF, Lichtman MA. Marrow cell egress. The central interaction of barrier pore size and cell maturation. J Clin Invest 1973;52:1154.
256. Kovich NL, Lin N, Yednock T, et al. Stem cell factor modulates avidity of α4β1 and α5β1 integrins expressed on hematopoietic cell lines. Blood 1995;85: 159–167.
257. Rother K. Leucocyte mobilizing factor: a new biological activity derived from the third component of complement. Eur J Immunol 1972;2:550.
258. Terashima T, English D, et al. “Release of polymorphonuclear leukocytes from the bone marrow by interleukin-8.” Blood 1998;92(3):1062–1069.
259. Athens JW, et al. Leukokinetic studies. III. The distribution of granulocytes in the blood of normal subjects. J Clin Invest 1961;40:159.
260. Athens JW, et al. Leukokinetic studies. IV. The total blood, circulating and marginal granulocyte pools and the granulocyte turnover rate in normal subjects. J Clin Invest 1961;40:989.
261. Vejlens G. The distribution of leucocytes in the vascular system. Acta Pathol Microbiol Immunol Scand [Suppl] 1938;33:1.
262. Bishop CR, et al. Leukokinetic studies. XIII. A non-steady state kinetic evaluation of the mechanism of cortisol-induced granulocytosis. J Clin Invest 1968;47:249.
263. Toren M, Goffinet JA, Kaplow LS. Pulmonary bed sequestration of neutrophils during hemodialysis. Blood 1970;36:337–340.
264. Brubaker LH, Nolph KD. Mechanisms of recovery from neutropenia induced by hemodialysis. Blood 1971;38:623–631.
265. Craddock PR, Fehr J, Dalmasso AP, et al. Hemodialysis leukopenia: pulmonary vascular leukostasis resulting from complement activation by dialyzer cellophane membranes. J Clin Invest 1977;59:879–888.
266. Craddock PR, Hammerschmidt DE, White JG, et al. Complement (C5a)-induced granulocyte aggregation in vitro. A possible mechanism of complement-mediated leukostasis and leukopenia. J Clin Invest 1977;60:260–264.
267. Skubitz KM, Craddock PR. Reversal of hemodialysis granulocytopenia and pulmonary leukostasis. J Clin Invest 1981;67:1383–1391.
268. Craddock PR, Fehr J, Brigham KL, et al. Complement and leukocyte-mediated pulmonary dysfunction in hemodialysis. N Engl J Med 1977;769–774.
269. Hammerschmidt DE, Weaver LJ, Hudson LD, et al. Association of complement activation and elevated plasma-C5a with adult respiratory distress syndrome. Pathophysiological relevance and possible prognostic value. Lancet 1980;1: 947–949.
270. Alexanian R, Donohue DM. Neutrophilic granulocyte kinetics in normal man. J Appl Psychol 1965;20:803.
271. Athens JW, et al. Leukokinetic studies. X. Blood granulocyte kinetics in chronic myelocytic leukemia. J Clin Invest 1965;44:765.
272. Athens JW, et al. Leukokinetic studies. XI. Blood granulocyte kinetics in polycythemia vera, infection, and myelofibrosis. J Clin Invest 1965;44:778.
273. Galbraith PR, et al. Patterns of granulocyte kinetics in health, infection and in carcinoma. Blood 1965;25:683.
274. Craddock CG, et al. Evaluation of marrow granulocytic reserves in normal and disease states. Blood 1960;15:849; Am J Med 1960;28:711; In: Stohlman F Jr, ed. The kinetics of cellular proliferation. New York: Grune & Stratton, 1959; Blood 1959;14:50; Ser Haematol 1965;1:13.
275. Mauer AM, et al. An analysis of leukocyte radioactivity curves obtained with radioactive diisopropylfluorophosphate (DFP32). In: Stohlman F Jr, ed. The kinetics of cellular proliferation. New York: Grune & Stratton, 1959.
276. Meuret G, et al. Neutrophil kinetics in man. Studies using autotransfusion of 3H-DFP labeled blood cells and autoradiography. Blut 1972;26:97.
277. Duvall CP, Perry S. The use of 51-chromium in the study of leukocyte kinetics in chronic myelocytic leukemia. J Lab Clin Med 1968;71:614.
278. McCall MS, et al. The tagging of leukemic leukocytes with radioactive chromium and measurement of the in vivo cell survival. J Lab Clin Med 1955; 45:717.
279. McMillan R, Scott JL. Leukocyte labeling with 51chromium. I. Technic and results in normal subjects. Blood 1968;32:738.
280. Davidson JG, et al. The kinetics of radiochromate-labeled normal and leukemic lymphocytes. Clin Res 1969;17:150.
281. Eyre HJ, et al. Relative labeling of leukocytes, erythrocytes and platelets in human blood by 51chromium. Blood 1970;36:250.
282. McMillan R, et al. Radiochromate leukokinetics in chronic myelocytic leukemia (CML). Abstr XII Cong Intl Soc Hematol, 1968.
283. Scott JL, et al. Leukocyte labeling with 51chromium. IV. The kinetics of chronic lymphocytic leukemic lymphocytes. Blood 1973;41:155.
284. Spivak JL, Perry S. Evaluation of 51Cr as a leucocyte label. Br J Haematol 1973;25:321.
285. Vincent PC. The measurement of granulocyte kinetics. Br J Haematol 1977;36:1.
286. Dresch C, et al. Kinetic studies of 51Cr and DF32P-labelled granulocytes. Br J Haematol 1975;29:67.
287. McAfee JG, Thaker ML. Survey of radioactive agents for in vitro labeling of phagocytic leukocytes. I & II. J Nucl Med 1976;17:480–488.
288. Thakur ML, et al. Indium-111-labeled autologous leukocytes in man. J Nucl Med 1977;18:1014–1021.
289. Goodwin DA. Cell labeling with oxine chelates of radioactive metal ions. J Nucl Med 1978;19:557.
290. Gottlieb RA, Giesing HA, Zhu JY, et al. Cell acidification in apoptosis. G-CSF delays programmed cell death in neutrophils by upregulating the vacuolar H+-ATPase. Proc Natl Acad Sci U S A 1995;92:5965–5968.
291. Squier MK, Sehnert AJ, et al. Apoptosis in leukocytes. J Leukoc Biol 1995;57 (1):2–10.
292. Moulding DA, Quayle JA, et al. Mcl-1 expression in human neutrophils: regulation by cytokines and correlation with cell survival. Blood 1998;92(7): 2495–2502.
293. Gottlieb RA, Giesing HA, Engler RL, Babior BM. The acid deoxyribonuclease of neutrophils: a possible participant in apoptosis-associated genome destruction. Blood 1995;86:2414–2418.
294. Savill JS, Wyllie AH, Henson JE, et al. Macrophage; phagocytosis of aging neutrophils in inflammation. J Clin Invest 1989;83:865–875.
295. Savill JS, Henson PM, Haslett C. Phagocytosis of aged human neutrophils by macrophages is mediated by a novel “charge-sensitive” recognition mechanism. J Clin Invest 1989;84:1518–1527.
296. Savill J, Dransfield I, Hogg N, Haslett C. Vitronectin receptor-mediated phagocytosis of cells undergoing apoptosis. Nature 1990;343:170–173.
297. Inflammation. In: Keen WW, White JW, eds. An American text-book of surgery. Philadelphia: WB Saunders, 1892:10–25.
298. Liles WC, Dale DC, Klebanoff SJ. Glucocorticoids inhibit apoptosis of human neutrophils. Blood 1995;86:3181–3188.
299. Iwai K, Miyawaki T, Takizawa T, et al. Differential expression of bcl-2 and susceptibility to anti-fas-mediated cell death in peripheral blood lymphocytes, monocytes, and neutrophils. Blood 1994;84:1201–1208.
300. Mecklenburgh KI., Walmsley SR, et al. Involvement of a ferroprotein sensor in hypoxia-mediated inhibition of neutrophil apoptosis. Blood 2002;100(8): 3008–3016.
301. Molloy EJ, O’Neill AJ, et al. Sex-specific alterations in neutrophil apoptosis: the role of estradiol and progesterone. Blood 2003;102(7):2653–2659.
302. Tulchinsky D, Hobel CJ. Plasma human chorionic gonadotropin, estrone, estradiol, estriol, progesterone, and 17 alpha-hydroxyprogesterone in human pregnancy. 3. Early normal pregnancy. Am J Obstet Gynecol 1973;117(7): 884–893.
303. Maianski NA., Roos D, et al. Tumor necrosis factor alpha induces a caspase-independent death pathway in human neutrophils. Blood 2003;101(5):1987–1995.
304. Florey HW, Grant LH. Leucocyte migration from small blood vessels stimulated with ultraviolet light: an electron-microscope study. J Pathol 1961;82:13.
305. Marchesi VT, Florey HW. Electron micrographic observations on the emigration of leukocytes. Q J Exp Physiol 1960;458:343.
306. Boggs DR, et al. Leukokinetic studies. VIII. A search for an extramedullary tissue pool of neutrophilic granulocytes. Proc Soc Exp Biol Med 1964;115:792.
307. Isaacs R, Danielian AC. A study of the white blood corpuscles appearing in the saliva and their relation to those in the blood. Am J Med Sci 1927;174:70.
308. Klinkhamer JM. Quantitative evaluation of gingivitis and periodontal disease. I. The progranulocytic migratory rate. Periodontics 1968;6:297.
309. Addis T. The number of formed elements in the urinary sediment of normal individuals. J Clin Invest 1926;2:409.
310. Garnett ES. Observation on the kinetics of urinary granulocytic excretion. Clin Sci 1965;28:157.
311. Ambrus CM, Ambrus JL. Regulation of the leukocyte level. Ann N Y Acad Sci 1959;77:445.
312. Bierman HR, et al. The pulmonary circulation as a source of leukocytes and platelets in man. Science 1951;114:276.
313. Boggs DR, et al. Granulocytopoiesis in germfree mice. Proc Soc Exp Biol Med 1967;125:325.
314. Fliedner TM, et al. Über die Umsatzkinetik der Leukocyten bei keimfreien Mäusen. Schweiz Med Wochenschr 1966;96:1236.
315. Crosson FJ, et al. Neonatal sepsis at the Johns Hopkins Hospital, 1969–1975: bacterial isolates and clinical correlates. Johns Hopkins Med J 1977;140:37.
316. Eickhoff TC, et al. Neonatal sepsis and other infections due to group B streptococci. N Engl J Med 1964;271:1221.
317. Freedman RM, et al. A half century of neonatal sepsis at Yale: 1928–1978. Am J Dis Child 1981;135:140.
318. McCracken GH Jr, Shinefield HR. Changes in the pattern of neonatal septicemia and meningitis. Am J Dis Child 1966;112:33.
319. Overall JC Jr. Neonatal bacterial meningitis. J Pediatr 1970;76:499.
320. Ziai M, Haggerty RJ. Neonatal meningitis. N Engl J Med 1958;259:314.
321. Niswander KR, Gordon M. The women and their pregnancies. The collaborative perinatal study of the National Institute of Neurological Diseases and Stroke. Department of Health, Education and Welfare Publication No. (NIH) 73-379. Washington, DC: US Government Printing Office, 1972.
322. Klein JO, March SM. Bacterial sepsis and meningitis. In: Remminton JS, Klein JO, eds. Infectious diseases of the fetus and newborn infant. Philadelphia: WB Saunders, 1983.
323. Cates KL, Rowe JC, Ballow M. The premature infant as a compromised host. Curr Probl Pediatr 1983;13:1.
324. Lubchenco LO, et al. Intrauterine growth as estimated from liveborn birthweight data at 24 to 42 weeks of gestation. Pediatrics 1963;32:793.
325. Barak Y, et al. Granulocyte-macrophage colonies in culture of human fetal liver cells: morphologic and ultrastructural analysis of proliferation and differentiation. Exp Hematol 1980;8:837.
326. Kelemen E, Calvo W, Fliedner TM. Intravascular hemopoietic cells. In: Atlas of human hematopoietic development. Berlin: Springer-Verlag, 1979.
327. Thomas UB, Yoffey JM. Human fetal haematopoiesis. I. The cellular composition of foetal blood. Br J Haematol 1962;8:290.
328. Wheeler JG, et al. Neutrophil storage pool depletion in septic, neutropenic neonates. Pediatr Infect Dis 1984;3:407.
329. Ijima H, et al. Predominance of macrophage-colony formation in human cord blood. Exp Hematol 1982;10:324.
330. Chervenick PA, Boggs DR. In vitro growth of granulocytic and mononuclear cell colonies from blood of normal individuals. Blood 1971;37:131.
331. Christensen RD, et al. Granulocyte-macrophage progenitor cells in term and preterm neonates. J Pediatr 1986;109:1047.
332. Issaragrisil S. Correlation between hematopoietic progenitors and erythroblasts in cord blood. Am J Clin Pathol 1983;80:865.
333. Knudtzon S. In vitro growth of granulocytic colonies from circulating cells in human cord blood. Blood 1974;43:375.
334. Kurnick JE, Robinson WA. Colony growth of human peripheral white blood cells in vitro. Blood 1971;37:136.
335. McCredie KB, Hersh EM, Freireich EJ. Cells capable of colony formation in the peripheral blood of man. Science 1971;171:293.
336. Prindull G, Gabriel M, Prindull B. Circulating myelopoietic stem cells (CFUc). Blut 1981;43:109.
337. Prindull G, Prindull B, Meulen N. Haematopoietic stem cells (CFUc) in human cord blood. Acta Paediatr Scand 1978;129:413.
338. Christensen RD, Rothstein G. Pre- and postnatal development of granulocytic stem cells in the rat. Pediatr Res 1984;18:599.
339. Greenberg PF, Schrier SL. Granulopoiesis in neutropenic disorders. Blood 1973;41:753.
340. Richard KA, et al. Studies on the proliferative capacity of the in vitro colony-forming cell in human bone marrow. Aust N Z J Med 1973;3:361.
341. Tebbi K, et al. A comparison of granulopoiesis in culture from blood and marrow cells of nonleukemic individuals and patients with acute leukemia. Blood 1976;48:235.
342. Christensen RD, et al. Granulocytic stem cell (CFUc) proliferation in experimental group B streptococcal sepsis. Pediatr Res 1983;17:278.
343. Metcalf D. Hemopoietic colonies, recent results. In: Cancer research. New York: Springer-Verlag New York, 1977.
344. Christensen RD, et al. Blood and marrow neutrophils during experimental group B streptococcal infection: quantification of the stem cell, proliferative, storage and circulating pools. Pediatr Res 1982;16:549.
345. Erdman SH, et al. The supply and release of storage neutrophils; a development study. Biol Neonate 1982;41:132.
346. Christensen RD, et al. Circulating and storage neutrophil changes in experimental type II group B streptococcal sepsis. Pediatr Res 1980;14:806.
347. Christiansen RD. Treatment of experimental group B streptococcal infection with hybridoma antibody. Pediatr Res 1984;18:1093.
348. Christensen RD. The effect of hybridoma antibody administration upon neutrophil kinetics during experimental type III streptococcal sepsis. Pediatr Res 1983;17:795.
349. Christensen RD, Rothstein G. Efficiency of neutrophil migration in the neonate. Pediatr Res 1980;14:1147.
350. Christensen RD, et al. Granulocyte transfusion in septic canine neonates. Pediatr Res 1982;16:57.
351. Christensen RD, et al. The leukocyte left shift in clinical and experimental neonatal sepsis. J Pediatr 1981;98:101.
352. Harper TE, et al. Neonatal meningitis: measurement of neutrophil kinetics and evaluation of experimental treatments. Pediatr Res 1986;20:397.
353. Christensen RD, et al. Fatal early onset group B streptococcal sepsis with normal leukocyte counts. Pediatr Infect Dis 1985;4:242.
354. Zeligs BJ, et al. Age-dependent susceptibility of neonatal rats to group B streptococcal type III infection: correlation of severity of infection and response of myeloid pools. Infect Immun 1982;37:255.
355. Eitzman DV, Smith RT. The nonspecific inflammatory cycle in the neonatal infant. Am J Dis Child 1959;97:326.
356. Bullock JD, et al. Inflammatory response in the neonate re-examined. Pediatrics 1969;44:58.
357. Miller ME. Chemotactic function in the human neonate: humoral and cellular aspects. Pediatr Res 1971;5:487.
358. Boher A, Zeligs BJ, Bellanti JA. Chemotactic responses of various differentiational stages of neutrophils from human cord and adult blood. Infect Immun 1982;35:922.
359. Pahwa SG, et al. Cellular and humoral components of monocyte and neutrophil chemotaxis in cord blood. Pediatr Res 1977;11:677.
360. Tono-Oka T, et al. Characteristics of impaired chemotactic function in cord blood leukocytes. Pediatr Res 1979;13:148.
361. Klein RB, et al. Decreased mononuclear and polymorphonuclear chemotaxis in human newborns, infants, and young children. Pediatrics 1977;60:467.
362. Sacchi F, et al. Different maturation of neutrophil chemotaxis in term and preterm newborn infants. J Pediatr 1982;101:272.
363. Krause PJ, et al. Polymorphonuclear leukocyte adherence and chemotaxis in stressed and healthy neonates. Pediatr Res 1986;20:296.
364. Laurenti F, et al. Neutrophil chemotaxis in preterm infants with infections. J Pediatr 1980;96:468.
365. Schuit KE, Homisch L. Inefficient in vivo neutrophil migration in neonatal rats. J Leukoc Biol 1984;35:583.
366. Miller ME. Pathology of chemotaxis and random mobility. Semin Hematol 1975;12:59.
367. Mease AD, Burgess DP, Thomas FJ. Irreversible neutrophil aggregation: a mechanism of decreased newborn neutrophil chemotactic response. Am J Pathol 1981;104:98.
368. Olson TA, et al. Newborn polymorphonuclear leukocyte aggregation: a study of physical properties and ultrastructure using chemotactic peptides. Pediatr Res 1983;17:993.
369. Strauss RG, Snyder EL. Chemotactic peptide binding by intact neutrophils from human neonates. Pediatr Res 1984;18:63.
370. Anderson DC, Hughes BJ, Smith CW. Abnormal mobility of neonatal polymorphonuclear leukocytes: relation to impaired redistribution of surface adhesion sites by chemotactic factor or colchicine. J Clin Invest 1981;68:863.
371. Stark B, Kahane I. Cord blood polymorphonuclear leukocyte chemotaxis: differential response to formyl peptide and endotoxin-activated serum. Isr J Med Sci 1983;19:34.
372. Kimura GM, et al. Reduced concanavalin A capping of neonatal polymorphonuclear leukocytes (PMNs). Pediatr Res 1981;15:1271.
373. Mease AD, et al. Decreased phytohemagglutinin-induced aggregation and C5a-induced chemotaxis of human newborn neutrophils. Pediatr Res 1980;14:142.
374. Sacchi F, Hill HR. Defective membrane potential changes in neutrophils from human neonates. J Exp Med 1984;160:1247.
375. Strauss RG, Snyder EL. Uptake of extracellular calcium by neonatal neutrophils. J Leukoc Biol 1985;37:423.
376. Anderson DC, et al. Impaired motility of neonatal PMN leukocytes: relationship to abnormalities of cell orientation and assembly of microtubules in chemotactic gradients. J Leukoc Biol 1984;36:1.
377. Efrati P, Rozenszajn L. The morphology of buffy coat in normal human adults. Blood 1960;16:1012.
378. Allan RN, Alexander MK. A sex difference in the leucocyte count. J Clin Pathol 1968;21:691.
379. Gabbert WR, et al. The effect of age on the leukocyte count. Geriatrics 1947;2:96.
380. Olbrich O. Blood changes in the aged. Edinburgh Med J 1947;54:306.
381. Sturgis CC, Bethell FH. Quantitative and qualitative variations in normal leukocytes. Physiol Rev 1943;23:279; Med Scand 1958;160:99.
382. Ezeilo G. Non-genetic neutropenia in Africans. Lancet 1972;2:1003.
383. Casey AE. The diurnal levels of blood leukocytes in the normal rabbit. Proc Soc Exp Biol Med 1940;45:863.
384. Kennon BR, et al. A study of the white blood cell picture in six young men. Am J Physiol 1937;118:690.
385. Wachholder K, et al. Änderungen des weissen Blutbildes nach Nahrungasufnahme. Alimentäre Leukopenie und alimentäre Leukocytose. Pflugers Arch 1949;251:459.
386. Sharp GWG. The effect of light on diurnal leucocyte variation. J Endocrinol 1960;21:213.
387. Garrey WE, Bryan WR. Variations in white blood cell counts. Physiol Rev 1935;15:597.
388. Petersen WF, Berg M. Meteorological influences on leukocytic partition. Proc Soc Exp Biol Med 1933;30:832.
389. Kennedy WP, MacKay L. The normal leucocyte picture in a hot climate. J Physiol 1936;87:336.
390. Muchmore HG, et al. Neutropenia in healthy men at the south polar plateau. Arch Intern Med 1970;125:646.
391. Forwell GD. The response of the leukocyte count in man to environmental heat and to exercise. J Physiol 1954;124:66.
392. Cress CH, et al. The effect of anoxic and anemic anoxia on the leukocyte count. Am J Physiol 1943;140:299.
393. Verzar F. Die Zahl der Lymphocyten und eosinophilen Leukocyten in 1800 und 3450 m Höhe. Schweiz Med Wochenschr 1952;82:324.
394. Farris EJ. The blood picture of athletes as affected by intercollegiate sports. Am J Anat 1943;72:223.
395. Altschule MD, et al. Changes in leukocytes of the blood in man after electrically induced convulsions. Arch Neurol 1949;62:624.
396. Gabrilove JL, et al. The effect of the parenteral injection of epinephrine on leukocyte counts in normal subjects and in patients with Addison’s disease. Blood 1949;4:646.
397. Samuels AJ. Primary and secondary leucocyte changes following the intramuscular injection of epinephrine hydrochloride. J Clin Invest 1951;30:941.
398. Levine SA, Golden R. Some observations on paroxysmal rapid heart action with special reference to roentgen-ray measurements of the heart in and out of attacks. Arch Intern Med 1922;29:836.
399. Milhorat AT, et al. Leukocytosis during various emotional states. Arch Neurol Psych 1942;47:779.
400. Davis ME, Hult BE. Changes in circulating eosinophils in women during the menstrual cycle and reproduction. J Clin Endocrinol 1949;9:714.
401. Pepper H, Lindsay S. Levels of eosinophils, platelets, leukocytes and 17-hydroxycorticosteroids during normal menstrual cycle. Proc Soc Exp Biol Med 1960;104:145.
402. Nelson DH, et al. Blood levels of 17-hydroxycorticosteroids following the administration of adrenal steroids and their relation to levels of circulating leukocytes. J Clin Invest 1952;31:843.
403. John TJ. Leukocytosis during steroid therapy. Am J Dis Child 1966;111:68.
404. Gray AB, Telford RD, et al. Granulocyte activation induced by intense interval running. J Leukoc Biol 1993;53(5):591–597.
405. Leino L, Saarinen K, et al. Systemic suppression of human peripheral blood phagocytic leukocytes after whole-body UVB irradiation. J Leukoc Biol 1999;65(5):573–582.
406. Trentin JJ. Determination of bone marrow stem cell differentiation by stromal hemopoietic inductive microenvironments (HIM). Am J Pathol 1971;65: 621.
407. Cline MJ, et al. Organ interactions in the regulation of hematopoiesis: in vitro interactions of bone, thymus, and spleen with bone marrow stem cells in normal, S1/S1d and W/Wv mice. J Cell Physiol 1977;90:105.
408. Wolf NS. Dissecting the hematopoietic microenvironment. Stem cell lodgment and commitment, and the proliferation and differentiation of erythropoietic descendants in the S1/S1d mouse. Cell Tissue Kinet 1974;7:89; Reversibility of hematopoietic stem cell direction (not “commitment”) as influenced by the microenvironment. Blood Cells 1978;4:37.
409. Witte ON. Steel locus defines new multipotent growth factor. Cell 1990;63:5.
410. Dexter TM, et al. Conditions controlling the proliferation of haemopoietic stem cells in vitro. J Cell Physiol 1977;91:335.
411. Dexter TM, et al. Self-renewal of haemopoietic stem cells: the roles of the environment, of growth factors and of the src oncogene. Haematol Blood Transfus 1985;29:363.
412. Price TH, Chatta GS, et al. Effect of recombinant granulocyte colony-stimulating factor on neutrophil kinetics in normal young and elderly humans. Blood 1996;88(1):335–340.
413. Demetri GD, Griffin JD. Granulocyte colony-stimulating factor and its receptor. Blood 1991;78:2791–2808.
414. Gabrilove J. The development of granulocyte colony-stimulating factor in its various clinical applications. Blood 1992;80:1382–1385.
415. Metcalf D. The molecular biology and functions of the granulocyte-macrophage colony-stimulating factors. Blood 1986;67:257–267.
416. Rowe JM, Andersen JW, Mazza JJ, et al. A randomized placebo-controlled phase III study of granulocyte-macrophage colony-stimulating factor in adult patients (55 to 70 years of age) with acute myelogenous leukemia: a study of the Eastern Cooperative Oncology Group (E1490). Blood 1995;86:457–462.
417. Bunn P, Crowley J, Kelly K, et al. Chemoradiotherapy with or without granulocyte-macrophage colony-stimulating factor in the treatment of limited-stage small-cell lung cancer: a prospective phase III randomized study of the southwest oncology group. J Clin Oncol 1995;13:1632–1641.
418. Yuo A, Kitagawa S, Osaka A. Recombinant human granulocyte colony-stimulating factor as an activator of human granulocytes: potentiation of responses triggered by receptor mediate agonists and stimulation of C3bi receptor expression and adherence. Blood 1990;74:2114–2149.
419. Yong KL, Linch DC. Differential effects of granulocyte- and granulocyte-macrophage colony-stimulating factors (G- and GM-CSF) on neutrophil adhesion in vitro and in vivo. Eur J Haematol 1992;49:251–259.
420. Spertini O, Kansas GS, Munro JM, et al. Regulation of leukocyte migration by activation of the leukocyte adhesion molecule-1 (LAM-1) selectin. Nature 1991; 349:691–694.
421. Ohsaka A, Kitagawa S, Sakamoto S, et al. In vivo activation of human neutrophil functions by administration of recombinant human granulocyte colony stimulating factor in patients with malignant lymphoma. Blood 1989;74: 2743–2748.
422. Avalos B, Gassos JC, Hedvat C, et al. Human granulocyte colony-stimulating factor: biologic activities and receptor characterization on hematopoietic cells and small cell lung cancer cell lines. Blood 1990;75:851–857.
423. Balazovich KJ, Almeida HI, Boxer LA. Recombinant human G-CSF and GM-CSF prime human neutrophils for superoxide production through different signal transduction mechanisms. J Lab Clin Med 1991;118:576–584.
424. Khwaja A, Carver JE, Linch DC. Interactions of granulocyte-macrophage colony-stimulating factor (CSF), granulocyte CSF, and tumor necrosis factor α in the priming of the neutrophil respiratory burst. Blood 1992;79:745–753.
425. Morstyn G, Souza LM, Keech J, et al. Effect of granulocyte colony-stimulating factor on neutropenia induced by cytotoxic chemotherapy. Lancet 1988;1: 667–672.
426. Lindemann A, Herrmann F, Oster F, et al. Hematologic effects of recombinant granulocyte colony-stimulating factor in patients with malignancy. Blood 1989;74:2644–2651.
427. de Haas M, Kerst JM, van der Schoot CE, et al. Granulocyte colony-stimulating factor administration to healthy volunteers: analysis of the immediate activating effects on circulating neutrophils. Blood 1994;84:3885–3894.
428. Berkow RL, Wang D, Larrick JW, et al. Enhancement of neutrophil superoxide production by preincubation with recombinant human tumor necrosis factor. J Immunol 1987;139:3783–3791.
429. Borish L, Rosenbaum R, Albury L, Clark S. Activation of neutrophils by recombinant IL-6. Cell Immunol 1989;121:280–289.
430. Sullivan GW, Carper HT, Sullivan JA, et al. Both recombinant interleukin-1 (beta) and purified human monocyte interleukin-1 prime human neutrophils for increased oxidative activity and promote neutrophils spreading. J Leukoc Biol 1989;45:389–395.
431. Yuo A, Kitagawa S, Kasahara T, et al. Stimulation and priming of human neutrophils by interleukin-8: cooperation with tumor necrosis factor and colony-stimulating factors. Blood 1991;78:2708–2714.
432. Lord BI, et al. A stimulator of stem cell proliferation in regenerating bone marrow. Biomedicine 1977;27:223.
433. Broxmeyer HE, et al. Cell-free granulocyte colony inhibiting activity derived from human polymorphonuclear neutrophils. Exp Hematol 1977;5:87; Science 1978;199:552.
434. Oshita AK, et al. cGMP stimulation of stem cell proliferation. Blood 1977; 49:585.
435. Broxmeyer HE, Lu L. Control of myelopoietic growth factor production. In: Progr Clin Biol Res 1985;184:145–155.
436. Heaney ML, Golde DW. Soluble cytokine receptors. Blood 1996;87:847–857.
437. Fliedner TM, et al. Morphologic and cytokinetic aspects of bone marrow stroma. In: Stohlman F Jr, ed. Hemopoietic cellular proliferation. New York: Grune & Stratton, 1970.
438. Katz R, et al. Studies of the purification of the leukocytosis-inducing factor (LIF). J Reticuloendothel Soc 1966;3:103.
439. Dornfest BS, et al. Mechanisms of leukocyte production and release. II. Factors influencing release from isolated perfused rat legs. J Lab Clin Med 1962;60:777.
440. Boggs DR, et al. Mechanisms controlling homeostasis of neutrophilic leukocytes. Haematol Lat 1967;10:43.
441. Boggs DR, et al. Neutrophilia-induced activity in plasma of dogs recovering from drug-induced myelotoxicity. Am J Physiol 1966;211:51.
442. Handler ES, et al. Mechanisms of leukocyte production and release. V. Studies on the leukocytosis-inducing factor in the plasma of rats treated with typhoid-paratyphoid vaccine. J Lab Clin Med 1966;67:398.
443. Boggs DR, et al. Neutrophil releasing activity in plasma of normal human subjects injected with endotoxin. Proc Soc Exp Biol Med 1968;127:689.
444. Lichtman MA, et al. The regulation of the release of granulocytes from normal marrow. In: Greenwelt TJ, Jamieson GA, eds. The granulocyte: function and clinical utilization. New York: Alan R. Liss, 1977.
445. Hunter J. A treatise on the blood, inflammation and gun-shot wounds. Nicol, London, 1774.
446. Hammerschmidt DE, Harris PD, Wayland H, et al. Complement-induced granulocyte aggregation in vivo. Am J Pathol 1981;102:146–150.
447. Gerard NP, Gerard C. The chemotactic receptor for human C5a anaphylatoxin. Nature 1991;349:614–617.
448. von Tscharner V, Deranleau DA, Baggiolini M. Calcium fluxes and calcium buffering in human neutrophils. J Biol Chem 1986;261:10163–10168.
449. Krause KH, Lew PD. Subcellular distribution of Ca2+ pumping sites in human neutrophils. J Clin Invest 1987;80:107–116.
450. Theler JM, Lew DP, Jaconi ME, et al. Intracellular pattern of cytosolic Ca2+ changes during adhesion and multiple phagocytosis in human neutrophils. Dynamics of intracellular Ca2+ stores. Blood 1995;85:2194–2201.
451. Barrowman MM, Cockcroft S, Gomperts BD. Differential control of azurophilic and specific granule exocytosis in Sendai-virus-permeabilized rabbit neutrophils. J Physiol 1987;383:115.
452. Lew PD, Monod A, Waldvogel FA, et al. Quantitative analysis of the cytosolic free calcium deficiency of exocytosis from three subcellular compartments in intact human neutrophils. J Cell Biol 1986;102:2197.
453. Smolen JE, Stoehr SJ, Boxer LA. Human neutrophils permeabilized with digitonin respond with lysosomal enzyme release when exposed to micromolar levels of free calcium. Biochem Biophys Acta 1986;886:1–17.
454. Smolen JE, Korchak HM, Weissmann G. The roles of extracellular and intracellular calcium in lysosomal enzyme release and superoxided anion generation by human polymorphonuclear leukocytes. Biochim Biophys Acta 1981; 677:512.
455. Lew PD, Andersson T, Hed J, et al. Ca2+-dependent and Ca2+-independent phagocytosis in human neutrophils. Nature 1985;315:509.
456. Balazovich KJ, Smolen JE, Boxer LA. Endogenous inhibitor of protein kinase C: association with human peripheral blood neutrophils but not with specific granule-deficient neutrophils or cytoplasts. J Immunol 1986;137:1665.
457. Cui Y, Harvey K, Akard L, et al. Regulation of neutrophil responses by phosphotyrosine phosphatase. J Immunol 1994;152:5420–5428.
458. Bokoch GM. Chemoattractant signaling and leukocyte activation. Blood 1995; 86:1649–1660.
459. Lowell CA, Berton G. Integrin signal transduction in myeloid leukocytes. J Leukoc Biol 1999;65(3):313–320.
460. Cronstein BN, Levin RI, Belanoff J, et al. Adenosine: an endogenous inhibitor of neutrophil-mediated injury to endothelial cells. J Clin Invest 1986;78:760–770.
461. Cronstein BN, Levin RI, Philips M, et al. Neutrophil adherence to endothelium is enhanced via adenosine A1-receptors and inhibited by adenosine A2-receptors. J Immunol 1992;148:2201–2206.
462. Skubitz KM, Wickham NW, Hammerschmidt DE. Endogenous and exogenous adenosine inhibit granulocyte aggregation without altering the associated rise in intracellular calcium concentration. Blood 1988;72:29–33.
463. Condliffe AM, Chilvers ER, et al. Priming differentially regulates neutrophil adhesion molecule expression/function. Immunology 1996;89(1):105–111.
464. Brandolini L, Bertini R, et al. IL-1 beta primes IL-8-activated human neutrophils for elastase release, phospholipase D activity, and calcium flux. J Leukoc Biol 1996;59(3):427–434.
465. Roberts PJ, Williams SL, et al. The regulation of neutrophil phospholipase A2 by granulocyte-macrophage colony-stimulating factor and its role in priming superoxide production. Br J Haematol 1996;92(4):804–814.
466. Mansfield PJ, Hinkovska-Galcheva V, et al. Granulocyte colony-stimulating factor primes NADPH oxidase in neutrophils through translocation of cytochrome b(558) by gelatinase-granule release. J Lab Clin Med 2002;140 (1):9–16.
467. Brown GE, Reiff J, et al. Maintenance and down-regulation of primed neutrophil chemiluminescence activity in human whole blood. J Leukoc Biol 1997; 62(6):837–844.
468. Ward PA, Becker EL. The deactivation of rabbit neutrophils by chemotactic factor and the nature of the activatable esterase. J Exp Med 1968;127(4):693–709.
469. O’Flaherty JT, Kreutzer DL, et al. Selective neutrophil desensitization to chemotactic factors. J Cell Biol 1979;80(3):564–572.
470. Webster RO, Hong SR, et al. Biologial effects of the human complement fragments C5a and C5ades Arg on neutrophil function. Immunopharmacology 1980;2(3):201–219.
471. Genco RJ, Van Dyke TE, et al. 1985 Kreshover lecture. Molecular factors influencing neutrophil defects in periodontal disease. J Dent Res 1986;65(12): 1379–1391.
472. Skubitz KM, Craddock PR, et al. Corticosteroids block binding of chemotactic peptide to its receptor on granulocytes and cause disaggregation of granulocyte aggregates in vitro. J Clin Invest 198168(1):13–20.
473. Walther A, Riehemann K, et al. A novel ligand of the formyl peptide receptor: annexin I regulates neutrophil extravasation by interacting with the FPR. Mol Cell 2000;5(5):831–840.
474. Hayhoe RP, Kamal AM, et al. Annexin 1 and its bioactive peptide inhibit neutrophil-endothelium interactions under flow: indication of distinct receptor involvement. Blood 2006;107(5):2123–2130.
475. Chadzinska M, Kolaczkowska E, et al. Morphine modulation of peritoneal inflammation in Atlantic salmon and CB6 mice. J Leukoc Biol 1999;65(5): 590–596.
476. Blixt A, Johnson P, Braide M, Bagge U. Microscopic studies on the influence of erythrocyte concentration on the post-junctional radial distribution of leukocytes at small venular bifurcations. Int J Microcirc Clin Exp 1985;4:141–156.
477. Schmid-Schonbein GW, Usami S, Skalak R, Chien S. The interaction of leukocytes and erythrocytes in capillary and postcapillary vessels. Microvasc Res 1980;19:45–70.
478. Metchnikoff E. In: Lectures on the comparative pathology of inflammation. New York: Dover, 1968:135–137.
479. Springer TA. Traffic signals for lymphocyte recirculation and leukocyte emigration: the multistep paradigm. Cell 1994;76:301–314.
480. Alon R, Hammer DA, Springer TA. Lifetime of the P-selectin-carbohydrate bond and its response to tensile force in hydrodynamic flow. Nature 1995;374: 539–542.
481. Carman CV, Springer TA. A transmigratory cup in leukocyte diapedesis both through individual vascular endothelial cells and between them. J Cell Biol 2004;167(2):377–388.
482. Kim M, Carman CV, Yang W, et al. The primacy of affinity over clustering in regulation of adhesiveness of the integrin αLβ2. J Cell Biol 2004;167(6): 1241–1253.
483. Senda N, et al. The mechanism of the movement of leucocytes. Exp Cell Res 1975;91:393.
484. Boyden SV. The chemotactic effect of mixtures of antibody and antigen on polymorphonuclear leucocytes. J Exp Med 1962;115:453.
485. Baum J, et al. A simplified method for the measurement of chemotaxis of polymorphonuclear leukocytes from human blood. J Lab Clin Med 1971;77:501.
486. de Duve C, Wattiaux R. Functions of lysosomes. Annu Rev Physiol 1966;28:435.
487. Metchnikoff E. Sur la lutte des cellules de l’organisme contre l’invasion des microbes. Ann Inst Pasteur Microbiol 1887;1:321.
488. Wright DG, Gallin JI. A functional differentiation of human neutrophil granules: generation of C5a by a specific (secondary) granule product and inactivation of C5a by azurophil (primary) granule products. J Immunol 1977;119:1068–1076.
489. Babior BM. The respiratory burst of phagocytes. J Clin Invest 1984;73:599.
490. Klebanoff SJ. Antimicrobial mechanisms in neutrophilic polymorphonuclear leukocytes. Semin Hematol 1975;12:117; In: Bellanti JA, Dayton DH, eds. The phagocytic cell in host resistance. New York: Raven Press, 1975.
491. Baldridge CW, Gerard RW. The extra respiration of phagocytosis. Am J Physiol 1933;103:235–236.
492. Rossi F, Zatti M. Biochemical aspects of phagocytosis in polymorphonuclear leucocytes. NADH and NADPH oxidation by the granules of resting and phagocytizing cells. Experientia 1964;20(1):21–23.
493. Babior BM, Kipnes RS, et al. Biological defense mechanisms. The production by leukocytes of superoxide, a potential bactericidal agent. J Clin Invest 1973;52 (3):741–744.
494. Hampton MB, Kettle AJ, et al. Inside the neutrophil phagosome: oxidants, myeloperoxidase, and bacterial killing. Blood 1998;92(9):3007–3017.
495. DeLeo FR, Quinn MT. Assembly of the phagocyte NADPH oxidase: molecular interaction of oxidase proteins. J Leukoc Biol 1996;60(6):677–691.
496. Curnette JT. Chronic granulomatous disease: the solving of a clinic riddle at the molecular level. Clin Immunol Immunopathol 1993;67:S2–S15.
497. Babior BM. NADPH oxidase. Curr Opin Immunol 2004;16(1):42–47.
498. Nauseef WM. Assembly of the phagocyte NADPH oxidase. Histochem Cell Biol 2004;122(4):277–291.
499. Klebanoff SJ. Myeloperoxidase: friend and foe. J Leukoc Biol 2005; 77(5): 598–625.
500. Klebanoff SJ. Myeloperoxidase: contribution to the microbicidal activity of intact leukocytes. Science 1970;169:1095.
501. Klebanoff SJ, White LR. Iodination defect in the leukocytes of a patient with chronic granulomatous disease of childhood. N Engl J Med 1969;280:460.
502. Test ST, Lampert MB, Ossanna PJ, et al. Generation of nitrogen-chlorine oxidants by human phagocytes. J Clin Invest 1984;74:1341–1349.
503. Malawista SE, Montgomery RR, Van Blaricom G. Evidence for reactive nitrogen intermediates in killing of staphylococci by human neutrophil cytoplasts. A new microbicidal pathway for polymorphonuclear leukocytes. J Clin Invest 1992;90: 631–636.
504. Maly FE, Schurer-Maly CC. How and why cells make superoxide: the “phagocytic” NADPH oxidase. NIPS 1995;10:233–238.
505. Lehrer RI, Cline MJ. Leukocyte myeloperoxidase deficiency and disseminated candidiasis. J Clin Invest 1969;48:1478.
506. Babior BM. Oxidants from phagocytes: agents of defense and destruction. Blood 1984;64:959.
507. Lakshmi VM, Nauseef WM, et al. Myeloperoxidase potentiates nitric oxide-mediated nitrosation. J Biol Chem 2005;280(3):1746–1753.
508. Eiserich JP, Hristova M, et al. Formation of nitric oxide-derived inflammatory oxidants by myeloperoxidase in neutrophils. Nature 1998;391(6665):393–397.
509. Nath J, Powledge A. Modulation of human neutrophil inflammatory responses by nitric oxide: studies in unprimed and LPS-primed cells. J Leukoc Biol 1997;62(6):805–816.
510. Evans TJ, Buttery LD, et al. Cytokine-treated human neutrophils contain inducible nitric oxide synthase that produces nitration of ingested bacteria. Proc Natl Acad Sci U S A 1996;93(18):9553–9558.
511. Weinberg JB. Nitric oxide production and nitric oxide synthase type 2 expression by human mononuclear phagocytes: a review. Mol Med 1998;4(9): 557–591.
512. Wheeler MA, Smith SD, et al. Bacterial infection induces nitric oxide synthase in human neutrophils. J Clin Invest 1997;99(1):110–116.
513. Jensen MS, Bainton DF. Temporal changes in pH within the phagocytic vacuole of the polymorphonuclear neutrophilic leukocyte. J Cell Biol 1973;56:379.
514. Mandell GL. Intraphagosomal pH of human polymorphonuclear neutrophils. Proc Soc Exp Biol Med 1970;134:447.
515. Rous PJ. On the mobilization of acid material within cells. J Exp Med 1925; 41:399.
516. Padgett GA, Hirsch JG. Lysozyme: its absence in tears and leukocytes of cattle. Aust J Exp Biol Med Sci 1967;45:569.
517. Baggilioni M, et al. Association of lactoferrin with specific granules in rabbit heterophil leukocytes. J Exp Med 1970;131:559.
518. Spitznagel JK. Nonoxidative antimicrobial reactions of leukocytes. In: Snyderman R, ed. Contemporary topics in immunobiology, vol 14. New York: Plenum, 1984.
519. Masson PL, et al. Lactoferrin, an iron-binding protein in neutrophilic leukocytes. J Exp Med 1969;130:643.
520. Arnold RR, et al. A bactericidal effect for human lactoferrin. Science 1977;197:263.
521. Martin E, Ganz T, et al. Defensins and other endogenous peptide antibiotics of vertebrates. J Leukoc Biol 1995;58(2):128–136.
522. Baggilioni M, et al. Further biochemical and morphological studies of granule fractions from rabbit heterophil leukocytes. J Cell Biol 1970;45:586.
523. Brewer DB. Electron microscopy of phagocytosis of staphylococci. J Pathol 1963;86:299.
524. Owen CA, Campbell EJ. The cell biology of leukocyte-mediated proteolysis. J Leukoc Biol 1999;65(2):137–150.
525. Ayoub EM, Wannamaker LW. The fate of group A streptococci following phagocytosis. J Immunol 1967;99:1099.
526. Cohn ZA. The fate of bacteria within phagocytic cells. I. J Exp Med 1963; 117:27.
527. Rogers DE, Tompsett R. The survival of staphylococci within human leukocytes. J Exp Med 1952;95:209.
528. Weissman G. Effect on lysosomes of drugs useful in connective tissue disease. In: Campbell PN, ed. Interaction of drugs and subcellular components in animal cells. London: J & A Churchill, 1968.
529. Zucker-Franklin D. Electron microscope study of the degranulation of polymorphonuclear leukocytes following treatment with streptolysin. Am J Pathol 1965;47:419.
530. Baggiolini M, Dewald B. Exocytosis by neutrophils. In: Snyderman R, ed. Contemporary topics in immunobiology, vol 14. New York: Plenum, 1984.
531. Chikkappa G, et al. Correlation between various blood white cell pools and the serum B12–binding capacities. Blood 1971;37:142.
532. Wiernik P, Sepick AA. Clinical significance of serum and urinary muramidase activity in leukemia and other hematologic malignancies. Am J Med 1969; 46:330.
533. Lollike K, Kjeldsen L, Sengelov H, Borregaard N. Lysozyme in human neutrophils and plasma. A parameter of myelopoietic activity. Leukemia 1995;9:159–164.
534. Corcino J, et al. Release of vitamin B12–binding proteins by human leukocytes in vitro. J Clin Invest 1970;49:2250.
535. Stormorhen H. The release reaction of secretion. Scand J Haematol [Suppl] 1969;9:1.
536. Simons K, Weber T. The vitamin B12–binding protein in human leukocytes. Biochim Biophys Acta 1966;117:201.
537. Herbert V. Diagnostic and prognostic values of measurement of serum vitamin B12–binding proteins. Blood 1968;32:305.
538. Allen RH. Human vitamin B12 transport proteins. Prog Hematol 1975;9:57.
539. Osserman EF, Lawlor DF. Serum and urinary lysozyme in monocytic and monomyelocytic leukemia. J Exp Med 1966;124:921.
540. Fink ME, Finch SC. Serum neuramidase and granulocyte turnover. Proc Soc Exp Biol Med 1968;127:365.
541. Lord PC, Wilmoth LM, Mizel SB, McCall CE. Expression on interleukin 1 alpha and beta genes by human blood polymorphonuclear leukocytes. J Clin Invest 1991;87:1312–1321.
542. Lindemann A, Riedel D, Oster W, et al. Granulocyte-macrophage colony stimulating factor induces cytokine secretion by human polymorphonuclear leukocytes. J Clin Invest 1989;83:1308–1312.
543. Melani C, Mattia GF, Silvani A, et al. Interleukin-6 expression in human neutrophil and eosinophil peripheral blood granulocytes. Blood 1993;81: 2744–2749.
544. El Ouriaghli F, Fujiwara H, et al. Neutrophil elastase enzymatically antagonizes the in vitro action of G-CSF: implications for the regulation of granulopoiesis. Blood 2003;101(5):1752–1758.
545. Horwitz M, Benson KF, et al. Mutations in ELA2, encoding neutrophil elastase, define a 21-day biological clock in cyclic haematopoiesis. Nat Genet 1999;23(4):433–436.
546. Dale DC, Person RE, et al. Mutations in the gene encoding neutrophil elastase in congenital and cyclic neutropenia. Blood 2000;96(7):2317–2322.
547. Nassar T, Akkawi S, et al. Human alpha-defensin regulates smooth muscle cell contraction: a role for low-density lipoprotein receptor-related protein/alpha 2-macroglobulin receptor. Blood 2002;100(12):4026–4032.
548. Weiss SJ. Tissue destruction by neutrophils. N Engl J Med 1989;320:365–376.
549. Murrell GA, Francis MJ, et al. Modulation of fibroblast proliferation by oxygen free radicals. Biochem J 1990;265(3):659–665.
550. Stirpe F, Higgins T, et al. Stimulation by xanthine oxidase of 3T3 Swiss fibroblasts and human lymphocytes. Exp Cell Res 1991;192(2):635–638.
551. McCord JM. The evolution of free radicals and oxidative stress. Am J Med 2000;108(8):652–659.
552. Lalezari P, Nussbaum M, Gelman S, Spaet TH. Neonatal neutropenia due to maternal isoimmunization. Blood 1960;15:236–243.
553. Huizinga TWJ, Kleijer M, Tetteroo PAT, et al. Biallelic neutrophil NA-antigen system is associated with a polymorphism on the phospho-inositol-linked Fcγ-receptor III (CD16). Blood 1990;75:213–217.
554. Lalezari P, Murphy GB, Allen FH. NB1, a new neutrophil antigen involved in the pathogenesis of neonatal neutropenia. J Clin Invest 1971;50:1108–1115.
555. Stroncek DF, Skubitz KM, McCullough JJ. Biochemical characterization of the neutrophil-specific antigen NB1. Blood 1990;75:744–755.
556. Skubitz KM, Stroncek DF, Sun B. The neutrophil-specific antigen NB1 is anchored via a glycosyl-phosphatidylinositol linkage. J Leukoc Biol 1991;49: 163–171.
557. Goldschmeding R, van Dalen CM, Faber N, et al. Further characterization of the NB1 antigen as a variably expressed 56-62 kD GPI-linked glycoprotein of plasma membranes and specific granules of neutrophils. Br J Haematol 1992;81:336–345.
558. Bux J. Molecular nature of granulocyte antigens. Transfus Clin Biol 2001; 8(3):242–247.
559. Bux J. Nomenclature of granulocyte alloantigens. ISBT Working Party on Platelet and Granulocyte Serology, Granulocyte Antigen Working Party, International Society of Blood Transfusion. Transfusion 1999;39(6):662–663.
560. Salmon JE, Edberg JC, et al. Fc gamma receptor III on human neutrophils. Allelic variants have functionally distinct capacities. J Clin Invest 1990;85(4): 1287–1295.
561. Bredius RG, Fijen CA, et al. Role of neutrophil Fc gamma RIIa (CD32) and Fc gamma RIIIb (CD16) polymorphic forms in phagocytosis of human IgG1- and IgG3-opsonized bacteria and erythrocytes. Immunology 1994;83(4): 624–630.
562. Nagarajan S, Chesla S, et al. Ligand binding and phagocytosis by CD16 (Fc gamma receptor III) isoforms. Phagocytic signaling by associated zeta and gamma subunits in Chinese hamster ovary cells. J Biol Chem 1995;270(43): 25762–25770.
563. Goldschmeding R, van Dalen CM, et al. Further characterization of the NB 1 antigen as a variably expressed 56-62 kD GPI-linked glycoprotein of plasma membranes and specific granules of neutrophils. Br J Haematol 1992;81(3):336–345.
564. Matsuo K, Lin A, et al. Variations in the expression of granulocyte antigen NB1. Transfusion 2000;40(6):654–662.
565. Stroncek DF, Jaszcz W, et al. Expression of neutrophil antigens after 10 days of granulocyte-colony-stimulating factor. Transfusion 1998;38(7):663–668.
566. Chen SM, Dummler JS, Bakken JS, et al. Identification of a granulocytotrophic Ehrlichia species as the etiologic agent of human disease. J Clin Microbiol 1994;32:589–595.
567. Bakken JS, Dummler JS, Chen SM, et al. Human granulocytic ehrlichiosis in the upper Midwest United States: a new species emerging? JAMA 1994;272: 212–218.
568. Bakken JS, Kreuth J, Wilson-Nordskog C, et al. Clinical and laboratory characteristics of human granulocytic ehrlichiosis. JAMA 1996;275:199–205.
569. Goodman JL, Nelson C, Vitale B, et al. Direct cultivation of the causative agent of human granulocytic ehrlichiosis. N Engl J Med 1996;334:209–215.
570. McCaffrey RL, Allen LA. Pivotal Advance: Francisella tularensis LVS evades killing by human neutrophils via inhibition of the respiratory burst and phagosome escape. J Leuk Biol 2006;80:1224–1230.