35
I. OVERVIEW
The cells that make up the human body vary widely with respect to their metabolic and development rates. Thyroid hormones provide the brain with a global method to govern these processes outside of conditions of acute stress. The thyroid gland is located in the neck just below the larynx (Figure 35.1) and is regulated by the hypothalamus and the pituitary gland. The active hormones of the hypothalamic–pituitary–thyroid axis are triiodothyronine (T3) and thyroxine (T4). T3 and T4 induce transcription, translation, and synthesis of pumps, transporters, enzymes, and cellular scaffolding and contractile elements, thereby increasing the metabolic rate of peripheral tissues. The thyroid gland also has a minor role in Ca2+ homeostasis via parafollicular C cells. These cells release calcitonin, which decreases Ca2+ and PO43− levels through increased urinary excretion. The major regulators of Ca2+ homeostasis are the parathyroid glands located in the inferior and superior margins of the thyroid gland (see Figure 35.1) and vitamin D. Parathyroid hormone (PTH) increases circulating Ca2+ by stimulating bone resorption and increasing renal Ca2+ reabsorption. Vitamin D also increases circulating Ca2+ and PO43− levels by increasing absorption of these ions in the gastrointestinal (GI) tract. Ca2+ homeostasis is important for a host of cellular events, such as cell signaling (e.g., initiating muscle contraction and vesicle secretion) and sustaining action potentials (e.g., in cardiac myocytes) as well as maintaining bone mineral density.
II. HYPOTHALAMIC–PITUITARY–THYROID AXIS
Thyroid hormone secretion is regulated by a multitiered endocrine axis involving the hypothalamus and pituitary. Thyroid-stimulating hormone (TSH) released from the anterior pituitary increases production and secretion of T3and T4 from the thyroid gland.
A. Hypothalamus
Thyroid hormone secretion is regulated by three hypothalamic nuclei: the paraventricular nucleus, the arcuate nucleus, and the median eminence (Figure 35.2). Parvocellular neurons in these areas project to and secrete thyroid-releasing hormone (TRH) and somatostatin into the hypophyseal portal circulation. Both hormones target thyrotropes in the pituitary gland.
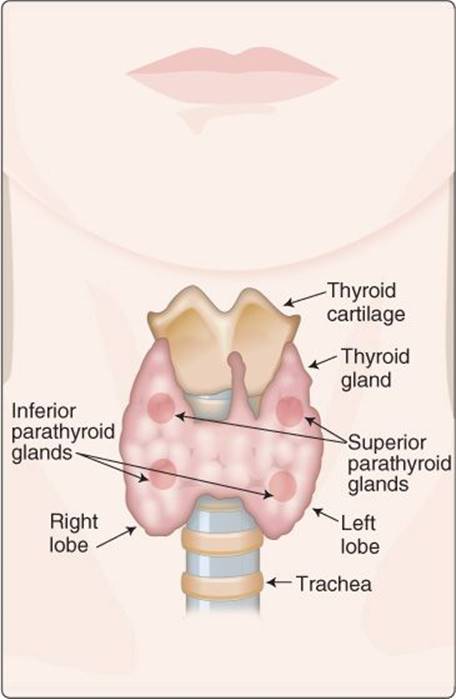
Figure 35.1
Thyroid and parathyroid glands.
B. Pituitary gland
TRH binds to thyrotrope TRH receptors, which are members of the G protein–coupled receptor (GPCR) superfamily that act through the phospholipase C second-messenger system (see 1·VII·B·3). TRH receptor occupancy stimulates TSH synthesis and release from secretory granules. In contrast, somatostatin (sometimes termed “somatotroph release–inhibiting factor”) decreases TSH production and release. Somatostatin binds to a different GPCR that inhibits adenylyl cyclase (AC) and cyclic adenosine monophosphate (cAMP) signaling (see 1·VII·B·2). Neurons within hypothalamic nuclei that initiate TSH release also express TSH receptors, which provides a negative feedback pathway by which high circulating TSH levels can inhibit further release.
C. Thyroid gland
TSH regulates the thyroid gland, which is an assemblage of numerous, 200 to 300-μm diameter hollow spheres (follicles) filled with a protein-rich fluid matrix known as colloid (Figure 35.3). Follicles are the site of thyroid hormone synthesis and secretion. Parafollicular C cells, which synthesize calcitonin, are randomly distributed between follicles throughout the gland. The TSH receptor is a GPCR that stimulates cAMP formation when occupied. cAMP, in turn, regulates most aspects of thyroid hormone secretion by regulating expression of most of the proteins involved in thyroid hormone synthesis (see Steps 1–8 in Section III·A).
1. Follicles: Follicles comprise a specialized epithelium composed of follicular cells resting on a basement membrane. The epithelium is supported by an extensive basolateral vasculature. The epithelium's apical side faces the colloid-filled follicular lumen. Follicular epithelial cells synthesize and secrete thyroid hormones when stimulated by TSH. TSH also increases expression of cellular components needed for thyroid hormone synthesis.
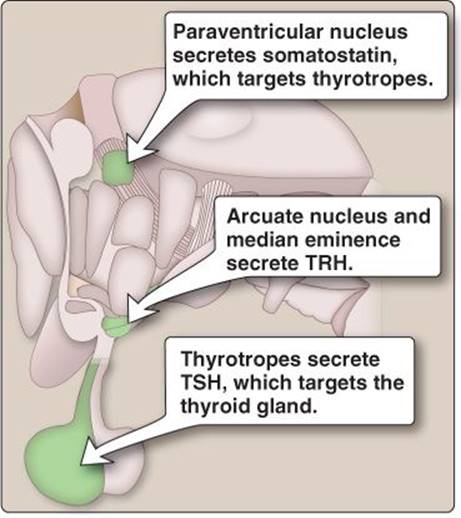
Figure 35.2
Hypothalamic nuclei involved in thyroid gland regulation. TRH = thyroid-releasing hormone; TSH = thyroid-stimulating hormone.
Clinical Application 35.1: Goiter
A normal thyroid gland weighs ~20 g, but its mass can increase quite dramatically under pathologic conditions. Enlarged thyroid glands (goiters) can be smooth or nodular depending on etiology. Thyroid cancers and infiltrates can cause nodular goiters. Goiters occur more commonly due to an excess production of thyroid-stimulating hormone (TSH) or activation of the TSH receptor. TSH receptor stimulation either by TSH or an autoimmune response causes gland growth (hypertrophy).
Goiter.
2. Colloid: The oxidative chemistry involved in thyroid hormone synthesis can be very harmful to cells, so it is performed extracellularly within colloid (see Figure 35.3). This is a similar concept to walling off hydrogen peroxide reactions within cytosolic peroxisomes. TSH increases the production of colloid.
III. THYROID HORMONES
The thyroid gland produces and secretes T3 and T4 in a ratio of ~1:10. T4 has a longer half-life than T3 but has a relatively low biologic activity. Conversion of T4 to T3 occurs primarily in target tissues. The thyroid hormones target virtually all cells in the body and exert their effects via cytosolic receptors that modulate gene expression.
A. Synthesis
Thyroid hormone synthesis and secretion is a multistep process involving iodination and conjugation of adjacent tyrosine residues (amino acid projections to which iodine can bind) on t hyroglobulin. Thyroglobulin's primary purpose is to hold thyroid hormone precursors in close proximity to enable the synthesis steps that occur in the colloid. Thyroid hormone synthesis begins with iodide up-take from the blood and can be broken into eight sequential steps (Figure 35.4).
1. Iodide uptake: Thyroid hormone synthesis begins with I− uptake (also called I− trapping) from the vasculature by follicular cells. I− is transported across the basolateral membrane from blood by a Na+-I−cotransporter (or, sodium/iodide symporter [NIS]), powered by the Na+ gradient established by the basolateral Na+-K+ ATPase.
Thyroid hormones cannot be produced without I−, which must be obtained from dietary sources (U.S. recommended dietary allowance is 150 g). Iodine deficiency results in hypothyroidism and presents as goiter (see Clinical Application 35.1).
2. Apical secretion: I− is then transported across the apical membrane primarily by a specialized Cl−-I− cotransporter known as a pendrin channel. Other mechanisms also exist for apical iodide transport, but, when this channel is defective, such as in Pendred syndrome, the patient presents with low circulating thyroid hormones. Thyroglobulin is synthesized in the follicular cells and is exocytosed across the apical membrane into the colloid.
3. Oxidation: The thyroglobulin-laden secretory vesicles express thyroid peroxidase (TPO), a heme-containing enzyme, on their inner surfaces. When the vesicles fuse with the apical membrane, TPO is presented to the colloid lumen and immediately catalyzes an oxidation reaction in which iodide is combined with H2O2 to form iodine (I2) and H2O. The H2O2 is derived from apical dual oxidase 2 (DUOX2) transporters that combine intermediates from the pentose phosphate pathway1 with O2 in the follicular cytosol to form H2O2 in the colloid.
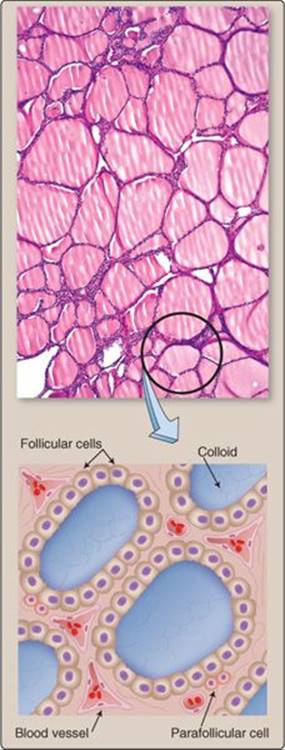
Figure 35.3
Cellular organization of the thyroid gland.
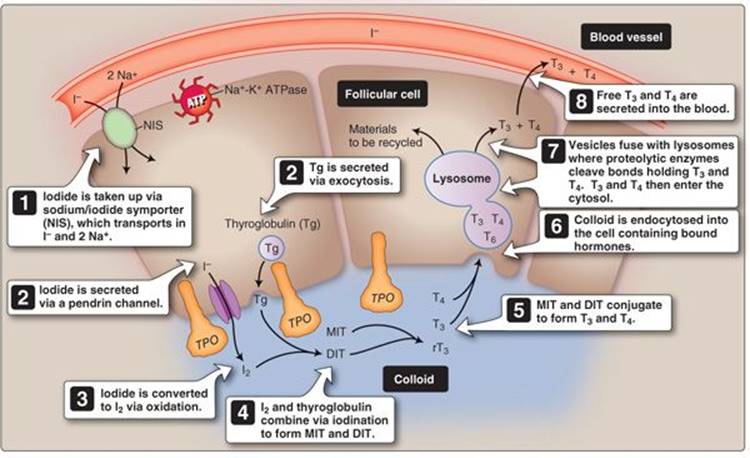
Figure 35.4
Thyroid hormone biosynthesis. ATP = adenosine triphosphate; DIT = diiodotyrosine; I− = iodide; I2 = iodine; MIT = monoiodotyrosine; T3 = triiodothyronine; T4 = thyroxine; TPO = thyroid peroxidase.
4. Iodination: TPO also facilitates iodination (or organification) of thyroglobulin tyrosine residues to form monoiodotyrosine (MIT) and diiodotyrosine (DIT) as shown in Figure 35.5. The precise reason for whether one or two iodines will bind to a particular residue is not clearly understood.
5. Conjugation: MIT and DIT combine (a process called conjugation) to form T3 and reverse T3 (rT3), whereas two DIT residues combine to form T4. The hormones remain attached to thyroglobulin until internalized by the follicular cells. Conjugation is also facilitated by TPO.2
 1For more information on the pentose phosphate pathway, see LIR Biochemistry, 5e, p. 145.
1For more information on the pentose phosphate pathway, see LIR Biochemistry, 5e, p. 145.
2Thyroid peroxidase can be inhibited pharmacologically by methimazole and propylthiouracil, which are used to treat a hyperfunctioning thyroid gland. See LIR Pharmacology, 5e, p. 298.
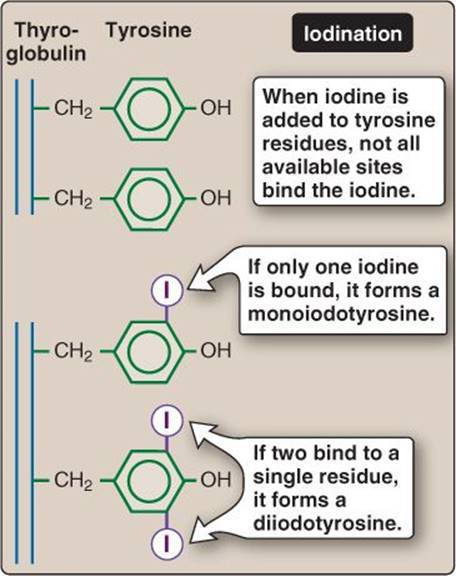
Figure 35.5
Thyroglobulin iodination.
6. Endocytosis: The iodinated and conjugated thyroglobulin is then endocytosed back into follicular cells, initiated by megalin receptors. TSH regulates megalin receptor expression and, thereby, indirectly controls the amount of colloid endocytosed.
7. Proteolysis: The endocytosed vesicle containing colloid then fuses with a lysosome, and the iodine-containing molecules are cleaved from thyroglobulin. T3 and T4 are released into the follicular cytosol near the basolateral membrane, and the remaining molecules and colloid material are recycled.
8. Secretion: The final step is secretion of T3 and T4 from the follicular cell into the blood. Cytosolic thyroid hormones diffuse through the basolateral cell membrane to the interstitial space, where they enter the capillary network and blood vessels of the highly vascularized thyroid gland.
B. Transport and regulation
Approximately 99% of the T3 and T4 released into the circulation binds to thyroid hormone-binding globulin and, to a lesser degree, albumin and transthyretin. Binding increases the half-life to as long as a week and serves as a thyroid hormone reserve for short periods. Both free (unbound) T3 and T4 participate in negative feedback control at the level of the hypothalamus and thyrotropes. In addition, thyroid hormones also increase somatostatin, which further decreases TSH release from thyrotropes (Figure 35.6).
C. Effects
T4 and T3 diffuse across the target cell membrane, and T4 is converted to T3 (the more biologically active form) by 5′- deiodinase (also called 5′/3′-monodeiodinase or deiodinase I). T3 then binds to a nuclear thyroid receptor that complexes with a retinoid X receptor (RXR). This receptor complex then binds to the thyroid-response element of DNA which, through both the addition of a coactivator and release of a corepressor, begins transcription (Figure 35.7). Proteins synthesized mediate a wide range of cellular responses, including increases in growth and development, glucose and fatty acid availability, and metabolic rate.
1. Growth and development: Growth and development of nervous tissue and bone are dependent on thyroid axis hormone synthesis and release. In nervous tissue, T3 and T4 aid in the timing and rate of development, which affects, for example, the development of stretch reflexes (see 11·III·B). In bone, thyroid hormone increases ossification and linear growth in children and adolescents. Thyroid hormone deficiencies can, thus, result in mental impairments and short stature in children.
2. Macronutrient metabolism: Thyroid hormones not only alter the rate of metabolism but also affect energy substrates. T3 and T4 increase both the breakdown of glycogen (glycogenolysis) and the formation of glucose (gluconeogenesis). T3 and T4 also increase the formation of lipids (lipogenesis) followed by promoting lipolytic enzymes, which break down the stored lipids into free fatty
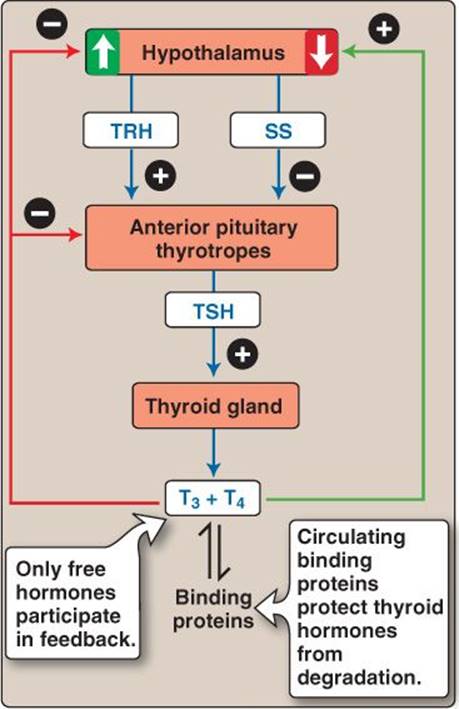
Figure 35.6
Feedback regulation of thyroid hormone release. SS = somatostatin; T3 = triiodothyronine; T4 = thyroxine; TRH = thyroid-releasing hormone; TSH = thyroid-stimulating hormone.
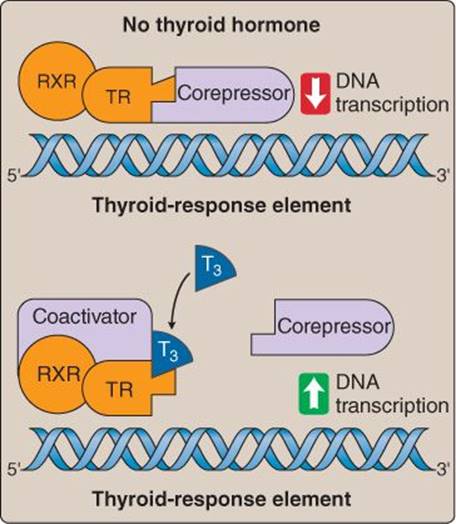
Figure 35.7
Thyroid hormone–receptor binding. RXR = retinoid X receptor; T3 = triiodothyronine; TR = thyroid hormone receptor.
Clinical Application 35.2: Hashimoto Disease
Hashimoto disease is the most common form of hypothyroidism, characterized by an autoimmune response that targets the thyroid gland. Inflammation and destruction of the gland decreases triiodothyronine and thyroxine production and release. A goiter may also form but is the result of the lymph and T-cell infiltrates rather than glandular hypertrophy. Patients lack energy, and are easily fatigued. They gain weight due to metabolic rate decline. If low levels of thyroid hormone occur during growth, growth can be stunted and development slowed.
3. Basal metabolic rate: Metabolism, through its inherent reaction inefficiencies, produces heat. Na+-K+ ATPase expression provides a good example of this phenomenon, in which thyroid hormone induces expression of the “always-on” pump uses more energy and produces more heat. The opposite occurs if pump expression is reduced by hypothyroid conditions (Figure 35.8).
4. Thyroid hormone and catecholamine synergy: When T3 and T4 and norepinephrine (from the sympathetic nervous system) are released in concert (e.g., during severe cold stress), the physiological functions of both are heightened. Thyroid hormones also upregu-late β-adrenergic receptors, which potentiates these synergistic effects.
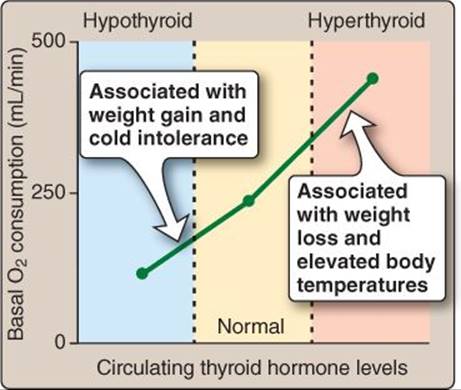
Figure 35.8
Thyroid hormone effects on metabolism.
IV. CALCITONIN
Calcitonin is a small (32 amino acid) peptide hormone produced by thyroid parafollicular C cells. Calcitonin is released in response to high plasma Ca2+ concentrations, and its primary effect is to block osteoclast-mediated bone resorption and Ca2+ mobilization (see 15·IV). Calcitonin is not thought to play any significant role in Ca2+ homeostasis in humans, but it can be used as a biomarker for thyroid cancer, and its physiologic effects can be exploited therapeutically.1 Plasma Ca2+ and PO43− levels are regulated primarily by PTH and vitamin D derivatives, as discussed below.
Clinical Application 35.3: Graves Disease
Graves disease produces hyperthyroidism via an antibody (thyroid-stimulating antibody) that mimics the effects of thyroid-stimulating hormone (TSH). TSH receptor stimulation causes glandular hypertrophy, which, in turn, causes goiter and other symptoms, such as myxedema and ophthalmopathy. Myxedema involves dermal edema and increases in connective tissue, which is caused by an overproduction of mucopolysaccharides (modified carbohydrates). Ophthalmopathy involves swelling around the eyes and also includes an excess production of mucopolysaccharides, which causes the eyes to protrude.
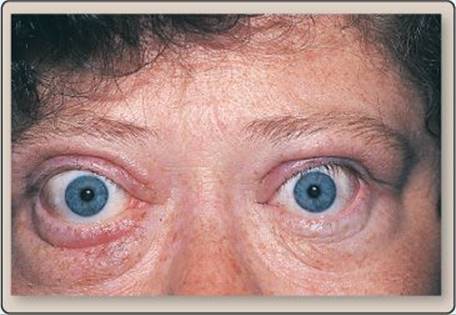
Exophthalmos.
V. PARATHYROID HORMONE AND VITAMIN D
PTH and vitamin D together regulate Ca2+ and PO43−. The major targets of regulation are the GI system (absorption), kidneys (reabsorption), and bone (deposition and resorption).
A. Calcium and phosphate pools
Large quantities of Ca2+ and PO43− are stored in bone (think of bone as a Ca2+ bank depository). This large Ca2+ source can be mobilized (withdrawn) during bone resorption in a process mediated by osteoclasts (see 15·IV). Alternately, Ca2+ can be actively stored (deposited) within bone during bone deposition in a process mediated by osteoblasts. Besides storage and release, Ca2+ balance is maintained by Ca2+ excretion from the kidney and Ca2+intake by the GI system (Figure 35.9). PO43− intake, excretion, and storage sites are similar to those of Ca2+.
B. Parathyroid hormone
PTH is released from parathyroid glands in response to a decline in circulating Ca2+ and Mg2+ levels. PTH actions are geared toward increasing Ca2+ availability.
1. Regulation: Parathyroid cells express a specialized GPCR that functions as a Ca2+ sensor. When plasma Ca2+ levels are high, the receptor tonically inhibits PTH secretion (Figure 35.10). The relationship between free Ca2+and PTH release is sigmoidal, with the steepest portion of the curve being in the physiologic range of plasma Ca2+. PTH release is similarly dependent on plasma free Mg2+.
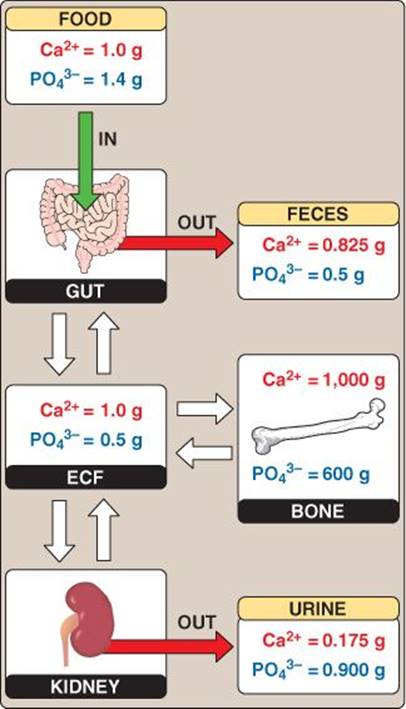
Figure 35.9
Calcium and phosphate balance. ECF = extracellular fluid.
Lithium sensitizes the Ca2+ receptor to changes in plasma Ca2+, causing increased PTH release in response to a given Ca2+ stimulus. This is the reason why bipolar patients on lithium salts for manic episodes can also have hypercalcemia.
1For a discussion of the use of calcitonin to treat osteoporosis, see LIR Pharmacology, 5e, p. 366.
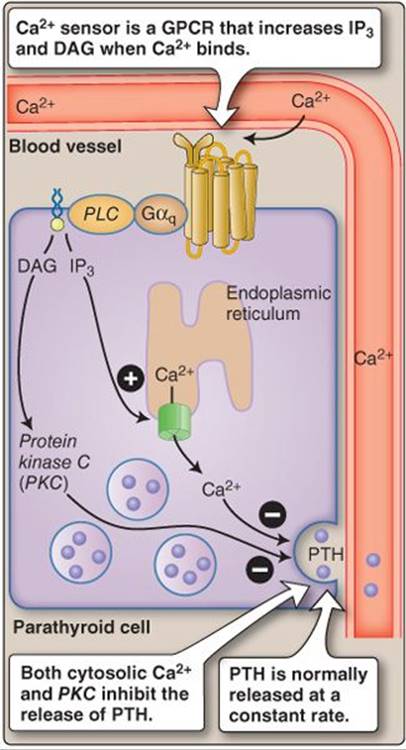
Figure 35.10
Calcium sensing. DAG = diacylglycerol; GPCR = G protein–coupled receptor; IP3 = inositol trisphosphate; PLC = phospholipase C; PTH = parathyroid hormone.
2. Function: PTH increases circulating Ca2+ levels by two principal mechanisms. First, it stimulates bone resorption by binding to receptors on the surface of osteoblasts, which then recruit osteoclasts precursors to a bone resorption site. Second, it stimulates Ca2+ reabsorption by the renal tubule (Figure 35.11). Proximal tubule reabsorption of PO43− is decreased by PTH. Although this increases PO43− excretion, it does not significantly alter circulating plasma PO43− levels because PTH also increases PO43− liberation from the bone. PTH also indirectly increases circulating Ca2+ by its stimulatory effects on 1α,25-dihydroxyvitamin D3 [1,25-(OH)2D3] synthesis.
C. Vitamin D derivatives
Vitamin D is functionally related to PTH, but, structurally, it is very different. Vitamin D and its derivatives are hydrophobic and transported in the blood primarily by vitamin D-binding protein. The primary active vitamin D derivative is 1,25-(OH)2D3, which increases Ca2+ and PO43− absorption by the small intestine. The synthesis of 1,25-(OH)2D3 from vitamin D2 or D3 involves a multistep process that includes both the liver and the kidney. The major regulatory enzyme for 1,25-(OH)2D3 synthesis is 25(OH)D1-α-hydroxylase. Vitamin D3 can be synthesized in the skin by keratinocytes (see 16·III·A·1) via interaction with UV light with 7-dehydrocholesterol, and vitamin D2 and D3 can also come from dietary sources.
1. Regulation: The primary factor that regulates 1,25-(OH)2D3 is PTH. PTH increases the activity of 25(OH)D1-α-hydroxylase and decreases the activity of 25(OH)D24-hydroxylase in the kidney. This shifts the reactions toward the production of 1,25-(OH)2D3. Low levels of both Ca2+ and PO43− as well as hormones, such as growth hormone, prolactin, and estrogen, also increase 1,25-(OH)2D3 levels.
2. Function: 1,25-(OH)2D3 is very effective in aiding the absorption of Ca2+ and PO43− from the GI tract and reabsorption of these ions from the renal tubule. Ca2+ absorption is improved by 1,25-(OH)2D3via increased synthesis of apical, basolateral, and cytosolic transport proteins. 1,25-(OH)2D3 binds to a nuclear vitamin D receptor that complexes with an RXR and induces transcription from the vitamin D response element. Thus, most of the effects of 1,25-(OH)2D3 are genomic, but there are some faster effects mediated by a cell membrane vitamin D receptor. These faster cell membrane responses are observed primarily in the GI system. PO43− absorption is also improved by 1,25-(OH)2D3 effects on apical Na+- PO4− cotransporter synthesis. 1,25-(OH)2D3 also aids in the maturation of osteoclasts, which potentially allows for more bone resorption. Finally, 1,25-(OH)2D3decreases PTH through negative feedback in addition to the negative feedback provided by increases in circulating Ca2+ due to increased intestinal absorption (Figure 35.12).
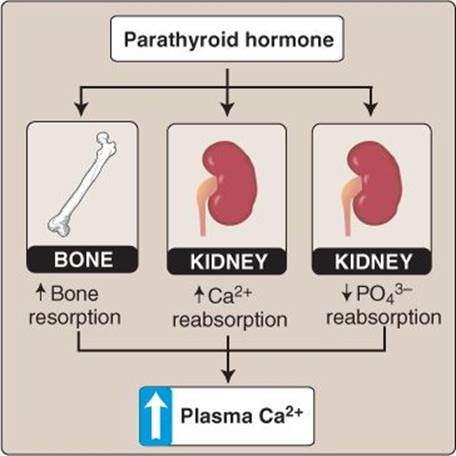
Figure 35.11
Parathyroid hormone effects.
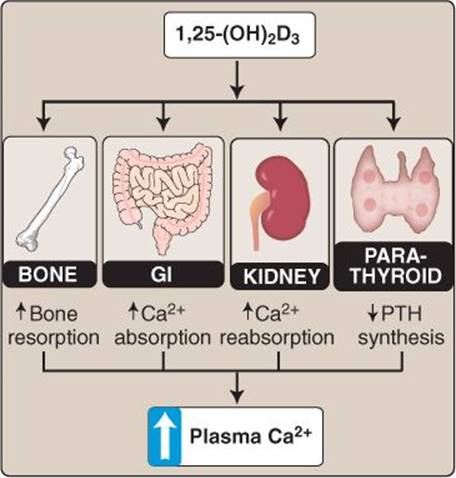
Figure 35.12
Vitamin D3. GI = gastrointestinal; PTH = parathyroid hormone.
Clinical Application 35.4: Rickets
Rickets is caused by a lack of vitamin D, Ca2+, or PO43−. Without adequate 1α,25-dihydroxyvitamin D3 [1,25-(OH)2D3], Ca2+ and PO43− cannot be absorbed from the gastrointestinal tract, causing plasma levels to fall. Parathyroid hormone is released as a result, promoting bone resorption to raise serum Ca2+ levels. Over time, bone minerals are depleted, causing them to weaken and bow. The lack of 1,25-(OH)2D3 can result from low exposure to sunlight, lack of access to dietary sources of vitamin D or a genetic disorder producing a low functioning 25(OH)D1-α-hydroxylase.
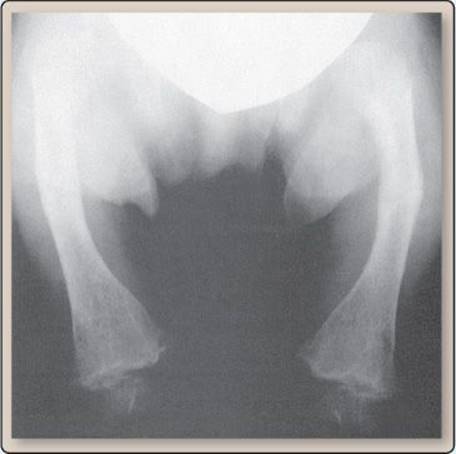
Bowed bone.
Chapter Summary
• Thyroid axis hormones consist of thyroxine (T4), triiodothyronine (T3), and reverse triiodothyronine. Thyroid intermediates consist of monoiodotyrosine (MIT) and diiodotyrosine (DIT). T4, which is formed from two DITs, is the most prevalent blood variant. T3 is the most biologically active in peripheral tissues and is formed from one DIT and one MIT.
• The steps of thyroid hormone synthesis are: uptake, colloid secretion, oxidation, iodination, conjugation, endocytosis, proteolysis, and glandular secretion. Thyroid-stimulating hormone (TSH) regulates the secretion of triiodothyronine and thyroxine via its actions on the Na+-I− symporter, pendrin channels, thyroid peroxidase, megalin receptors, and proteolytic enzymes. TSH also causes enlargement of the thyroid gland.
• The majority of triiodothyronine (T3) and thyroxine (T4) in blood is bound to thyroid hormone-binding globulin rendering them inactive, but some T3 and T4 circulates in the active free form.
• Once triiodothyronine (T3) and thyroxine (T4) enter a cell, the majority of the T4 is converted to T3, and T3 has a higher binding affinity for thyroid receptors. Thyroid hormones act via nuclear receptors, to modulate transcription and translation and, thereby, increase protein synthesis.
• Depending on which proteins are synthesized, thyroid hormones increase metabolic rate, heat production, and glycogen and fat breakdown and utilization. Thyroid hormones are also imperative for normal growth and development.
• Parathyroid hormone (PTH) increases circulating Ca2+ levels by increasing Ca2+ reabsorption in the kidney and by increasing bone resorption. PTH also activates vitamin D3.
• Vitamin D3 is converted to its active form via ultraviolet light exposure or enzymatic reaction in the kidney. The active form of vitamin D is 1α,25-dihydroxyvitamin D3 [1,25-(OH)2 D3], and its primary function is to increase circulating Ca2+ levels via absorption from the intestines, reabsorption from the kidney, and bone resorption.