The development of most of the organs and systems is touched upon in the text descriptions of the regions concerned. Here a very brief account of some important features of early development is included, to provide a background for the later notes.
Early development
For the first 8 weeks of the 40-week human gestation period the developing organism is an embryo; after that time it is a fetus. By the end of the embryonic period most organs have differentiated, and the changes during the fetal period are essentially those of maturation. Many but not all congenital defects are initiated in the embryo rather than the fetus.
The fertilized ovum or zygote undergoes repeated cell divisions (cleavage) to produce a mass of cells, the morula, which travels along the uterine tube towards the uterus. Further division enlarges the morula and a fluid-filled cavity (the extraembryonic coelom) appears in it; the whole structure is now a blastocyst. At this stage implantation into the uterine mucosa takes place, about 6 days after fertilization. The outer layer of cells of the blastocyst, the trophoblast, is destined to become placental. The remainder of the cells are concentrated at one end of the blastocyst to form the inner cell mass or embryoblast, attached to the inner aspect of the trophoblast.
At the beginning of the second week after fertilization, the embryoblast differentiates into two layers, a layer of columnar cells (the epiblast) and a layer of cuboidal cells (the hypoblast). Two cavities then appear, the amniotic cavity, which is related to the epiblast, and the yolk sac, which is related to the hypoblast (Fig. 1.16). The two cavities are surrounded by the extraembryonic coelom, except where the embryoblast is connected to the trophoblast by the connecting stalk, which later develops into the umbilical cord.
A primitive streak appears on the amniotic aspect of the embryoblast towards what will become the caudal end of the embryo (Fig. 1.17A). The streak indicates the site of a groove at the cephalic end of which is a primitive pit with slightly raised margins; this is the primitive node. In the third week, epiblast cells in the region of the primitive streak invaginate, displacing the hypoblast, and spread bilaterally and cephalad forming two new layers, the mesodermnext to the epiblast and the endoderm adjacent to the hypoblast. The remaining epiblast cells form the ectoderm. From the primitive pit a rod of cells invaginate directly cephalad between ectoderm and endoderm; this is the notochord, which extends as far as the buccopharyngeal membrane, where ectoderm and endoderm remain in contact. Similarly at the caudal end of the primitive streak, ectoderm and endoderm are in apposition at the cloacal membrane.
|
|
|
|
|
Figure 1.17 A Dorsal view of an embryo at the beginning of the third week; B formation of neural tube and neural crest. |
An indentation of the ectoderm overlying the notochord forms the neural groove (Fig. 1.17B). Its edges unite to form the neural tube, which becomes depressed below the surface. In due course, the brain and spinal cord develop from the neural tube. Some of the cells derived from the edges of the groove become isolated between the tube and the overlying ectoderm to form the neural crest. Its cells are destined to migrate and contribute to the development of several structures, including posterior root ganglia of spinal nerves, corresponding ganglia of cranial nerves, autonomic ganglia, neuroglia, Schwann cells, meninges, bones of the skull and face, sclera and choroid of the eye, dentine and cementum of teeth, parafollicular cells of the thyroid, chromaffin cells of the suprarenal medulla and melanocytes.
Alongside the notochord and neural tube the mesoderm lies in three longitudinal strips (Fig. 1.18). That nearest the midline is the paraxial mesoderm; it becomes segmented in cephalocaudal sequence into masses of cells called mesodermal somites. The somites produce: (1) the sclerotome, medially, which surrounds the neural tube and notochord, producing the vertebrae and ribs; and (2) the dermomyotome or muscle plate, laterally, which produces the muscles of the body wall and the dermis of the skin.
|
|
|
Figure 1.18 Cross-section through an embryo at the end of the third week. |
The intermediate strip of mesoderm is the intermediate cell mass. From its lateral side in cephalocaudal sequence develop successively the pronephros, mesonephros and metanephros and their associated ducts—the progenitors of the urinary and genital systems. Its medial side gives rise to the gonad and the cortex of the suprarenal gland.
The most lateral strip of mesoderm is the lateral plate. Very early the embryo begins to curl up, a result of the more rapid growth of the dorsal (ectodermal) surface. The embryo becomes markedly convex towards the amniotic cavity and correspondingly concave towards the yolk sac. As the lateral plate curls around to enclose the yolk sac its mesoderm becomes split into two layers by a space that appears within it. The space is the beginning of the intraembryonic coelom or body cavity. The inner layer is the splanchnic (visceral) mesoderm. It encloses the yolk sac in an hourglass constriction; the part of the yolk sac outside persists in the umbilical cord as the vitellointestinal duct; the part inside the embryo becomes the alimentary canal. The outer layer of the lateral plate is the somatic (parietal) mesoderm. Into it the paraxial myotomes migrate in segments to produce the flexor and extensor muscle layers of the body wall. The coelomic cavity at first includes pleural and peritoneal spaces in one continuum; they become separated later. The pleura and peritoneum are thus mesodermal in origin.
The limb buds grow from the lateral plate mesoderm and their muscles develop in situ. Although the lateral plate mesoderm is unsegmented, the motor fibres that grow into it from the spinal cord limb plexuses arrange their distribution in a segmental pattern.
The septum transversum consists of the mass of mesoderm lying on the cranial aspect of the coelomic cavity. Its cranial part contains the pericardial cavity, the walls of which develop into the pericardium and part of the diaphragm. It is invaded by muscles from cervical myotomes, mainly the fourth; they produce the muscle of the diaphragm. The septum transversum later descends, taking the heart with it, to the final position of the diaphragm. The caudal part of the septum transversum is invaded by the developing liver, which it surrounds as the ventral mesogastrium.
The folding of the embryo is impeded to some extent at the tail end by the presence of the connecting stalk, which later becomes the umbilical cord. The greatest amount of folding occurs at the head end of the embryo. By the end of the first fortnight the forebrain capsule is folded down over the pericardium, and a mouth pit, the stomodeum, shows as a dimple between the two. Within the body of the embryo the gut cavity extends headwards dorsal to the pericardium, as far forwards as the buccopharyngeal membrane, which closes the bottom of the mouth pit. The buccopharyngeal membrane breaks down and disappears in the fourth week and its former site cannot be made out with certainty in the later embryo or adult. Cranial to the site of the membrane the mouth pit is lined with ectoderm; this includes the region of all the mandibular and maxillary teeth, and the anterior two-thirds of the tongue. Rathke's pouch arises from this ectoderm and forms the anterior lobe of the pituitary gland. Caudal to the buccopharyngeal membrane is the pharynx, lined with endoderm and lying dorsal to the pericardium.
Pharyngeal arches and pouches
Mesodermal condensations develop in the side walls of the primitive pharynx to form the pharyngeal arches and they grow around towards each other ventrally, where they fuse in the midline. In this way a series of six horseshoe-shaped arches (also called branchial arches) comes to support the pharynx (Fig. 1.19). Deep grooves appear on the surface of the embryo at the intervals between the arches; these are the pharyngeal (or branchial) clefts. The fifth arch is rudimentary and only four clefts are visible. Outpouchings develop from the lining of the pharynx in between the arches and opposite the clefts: the pharyngeal (or branchial) pouches. The fourth and fifth pouches share a common opening into the lumen of the pharynx. In each arch a central bar of cartilage forms and muscle differentiates from the mesoderm around it. An artery and cranial nerve are allocated to the supply of each arch and its derivatives. Vascular patterns are very changeable during development, but a nerve supply, once established, remains constant and knowledge of the nerve supply of a muscle enables its pharyngeal arch origin to be determined.
|
|
|
Figure 1.19 Floor of the developing pharynx. The pharyngeal (branchial) arches are numbered. The foramen caecum lies in the midline between the first and second arches. |
First (mandibular) arch
Chondrification in the mesoderm of the first arch produces Meckel's cartilage. The dorsal end of Meckel's cartilage produces the incus and malleus, and the anterior ligament of the malleus. The sphenomandibular ligament (Fig. 1.20) is a remnant of the fibrous perichondrium of Meckel's cartilage. The lingula at the mandibular foramen develops from the cartilage. The mandible starts ossifying in membrane lateral to Meckel's cartilage, and the rest of the cartilage becomes incorporated in the developing mandible. Some time after birth the cartilage disappears.
|
|
|
Figure 1.20 Derivatives of the first arch cartilage. |
Ectodermal and endodermal derivatives of this arch are the mucous membrane and glands (but not the muscle) of the anterior two-thirds of the tongue. The muscles of mastication (masseter, temporal and pterygoids), the mylohyoid and anterior belly of digastric, and the two tensor muscles (tensor palati and tensor tympani) develop from the first arch and are supplied by the mandibular nerve, which is the nerve of this arch. Part of the artery of the first arch persists as the maxillary artery.
Other arches
The skeletal and muscular derivatives of the remaining arches can be summarized as follows.
Second (hyoid) arch
Skeletal derivatives: stapes; styloid process; stylohyoid ligament; lesser horn; and superior part of body of hyoid bone (Fig. 1.21).
|
|
|
Figure 1.21 Derivatives of the second and third arch cartilages. |
Muscular derivatives: muscles of facial expression (including buccinator and platysma); stapedius; stylohyoid; posterior belly of digastric—all supplied by the facial, the nerve of the second arch.
Third arch
Skeletal derivatives: greater horn and inferior part of body of hyoid bone (Fig. 1.21).
Muscular derivatives: stylopharyngeus—supplied by the glossopharyngeal, the nerve of the third arch.
Fourth and sixth arches
Skeletal derivatives: thyroid; cricoid; epiglottic and arytenoid cartilages.
Muscular derivatives: intrinsic muscles of larynx; muscles of pharynx; levator palati—all supplied by laryngeal and pharyngeal branches of the vagus, the nerve of these arches.
Lateral derivatives of the pharyngeal pouches
Except for the first, each pouch grows laterally into a dorsal and a ventral diverticulum.
First pouch. This is the only pouch in which the endoderm remains in close apposition to the ectoderm of the corresponding cleft, namely at the tympanic membrane, where the mesoderm separating them is minimal. In the other pouches the ectoderm and endoderm are finally widely separated. The endoderm of the first pouch is prolonged laterally, via the auditory tube, to form the middle ear and mastoid antrum (the second pouch gives a contribution to the middle ear; see below). The first pharyngeal cleft becomes deepened to form the external acoustic meatus.
Second pouch. The dorsal part assists the first pouch in the formation of the tympanic cavity, taking what may be its pretrematic nerve (the tympanic branch of the glossopharyngeal) with it; the word ‘trema’ means a cleft. The ventral part of the pouch develops the tonsillar crypts and the supratonsillar fossa from its endoderm, the surrounding mesoderm contributing the lymphatic tissue of the palatine tonsil. The nerve supply of these derivatives is the glossopharyngeal.
Third pouch. Dorsally the inferior parathyroid gland (termed parathyroid III) and ventrally the thymic rudiment grow from this pouch. The latter progresses caudally and joins with that of the other side to produce the bilobed thymus gland. In its descent the thymic bud draws parathyroid III in a caudal direction, so that ultimately the latter lies inferior to parathyroid IV, which is derived from the fourth pouch. From this thymic bud the medulla of the thymus, including the thymic (Hassall's) corpuscles, is derived; the lymphocytes of the cortex migrate from bone marrow.
Fourth pouch. The superior parathyroid glands (parathyroid IV) are derived from the endodermal lining of this pouch.
Fifth pouch. This forms the ultimobranchial body, from which are derived the parafollicular (C) cells of the thyroid gland which produce calcitonin.
Cervical sinus
Concurrently with the growth of the above derivatives from the endoderm of the pouches a change takes place externally in the overlying ectoderm. The only pharyngeal cleft to persist is the first, which forms the external ear. The second arch increases in thickness and grows caudally, over the third, fourth and sixth arches, covering the second, third and fourth clefts and meeting skin caudal to these. During this process a deep groove is formed, which becomes a deep pit, the cervical sinus. The lips of the pit then meet and fuse and the imprisoned ectoderm disappears. Persistence of this ectoderm gives rise to a branchial cyst. Persistence of the deep pit is termed a branchial sinus. A rare branchial fistula results from breaking down of the tissues between the floor of the pit and the side wall of the pharynx (endoderm). The track of the fistula runs from the region of the palatine tonsil, between the external and internal carotid arteries, and reaches the skin anterior to the lower end of sternocleidomastoid.
Ventral derivatives of the floor of the pharynx
The tongue, thyroid gland and larynx are derived from the floor of the mouth (Fig. 1.19).
Buds from the first, third and fourth arches form the stroma of the tongue, the epithelium being derived from the ectoderm of the stomodeum and the endoderm of the cranial end of the pharynx. Occipital myotomes migrate forwards to provide the musculature, carrying their nerve (hypoglossal) supply with them.
The thyroglossal duct originates from the endoderm of the floor of the pharynx at the foramen caecum (Fig. 1.19) in the region of the developing tongue, and then passes caudally in front of the hyoid bone, behind which it forms a recurrent loop (Fig. 1.22). The thyroid gland buds from the duct's distal end, which itself may give rise to the pyramidal lobe. Other remnants of the duct may persist as accessory thyroid glands or form thyroglossal cysts (Fig. 1.23). Rupture of a cyst gives rise to a thyroglossal sinus (‘fistula’). Removal of retrohyoid duct remnants usually requires excision of a segment from the middle of the hyoid bone. Failure of descent of the thyroglossal duct may result in the development of a lingual thyroid.
|
|
|
Figure 1.22 Course of the thyroglossal duct. The thyroid gland, larynx and trachea have been drawn to a smaller scale than the tongue, mandible and hyoid bone. |
|
|
|
Figure 1.23 Thyroglossal cyst. |
In the ventral wall of the pharynx a laryngotracheal groove appears. The cephalic end of this gutter is limited by the furcula, a ridge in the shape of a wish-bone (Fig. 1.19). The ridges which limit the gutter grow towards each other and, by their fusion, convert the gutter into a tube. While maintaining its connection with the pharynx at its cranial end, the larynx, the rest of the tube, the trachea, separates from the oesophagus and buds out into the bronchi and lungs at its caudal end. Failure of proper separation of the trachea and oesophagus results in a tracheo-oesophageal fistula. The furcula persists at the aperture of the larynx, whose cartilages, including that of the epiglottis, derive from the underlying fourth and sixth pharyngeal arches.
Branchial arch arteries
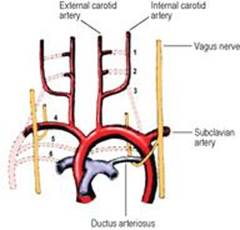
From the cephalic end of the primitive heart tube (see p. 30) a ventral aorta divides right and left into two branches which curve back caudally as the two dorsal aortae. As the branchial arches develop, a vessel in each arch joins the ventral to the dorsal aortae. Thus six aortic arches are to be accounted for. Caudal to this region, the two dorsal aortae fuse to become a single vessel; proximal to the fusion, a part of the right dorsal aorta subsequently disappears.
Parts of the first and second arch arteries form the maxillary and stapedial arteries respectively; the latter does not persist after birth. They are branches of the external carotid artery. The third remains as the common carotid and part of the internal carotid arteries. The fourth on the right contributes to the subclavian artery, on the left to the arch of the aorta. The fifth disappears entirely. By the time the sixth artery appears the upper bulbar part of the heart tube has been divided into aorta and pulmonary trunk and it is to the pulmonary trunk that the sixth arch arteries are connected ventrally. Dorsally they communicate with the dorsal aortae. The dorsal part of the sixth arch artery disappears on the right side but persists on the left as the ductus arteriosus, which thus connects the left pulmonary artery to the arch of the aorta (Fig. 1.24). This explains why the recurrent laryngeal (sixth arch) nerve hooks round the ligamentum arteriosum on the left, but migrates up and hooks round the subclavian artery on the right.
|
|
|
Figure 1.24 Fate of the arch arteries. The arteries are numbered and the dotted lines indicate the arteries that disappear. The asymmetry of the course of the recurrent laryngeal nerves results from differences in the fate of the lower arch arteries. |
Anomalies of the great vessels
The most common anomaly of development is a patent ductus arteriosus (persistence of part of the left sixth arch artery), which fails to close in the immediate postnatal period. Coarctation of the aorta (narrowing) is due to a defect of the tunica media which forms a shelf-like projection into the lumen, most commonly in the region of the ductus connection; collateral circulation distal to the obstruction is provided by the internal thoracic and posterior intercostal arteries. An abnormal origin of the right subclavian artery is from the arch of the aorta, just distal to the origin of the left one. The abnormal artery passes to the right behind the oesophagus and is a possible cause of dysphagia. With the lack of a normal right subclavian arch, the right recurrent laryngeal nerve is non-recurrent and runs down the side of the larynx: a possible hazard in thyroidectomy. The reported incidence of a non-recurrent nerve is around 1%.
Development of mouth and face
The stomodeum (mouth pit) has appeared by the end of the second week and the buccopharyngeal membrane between it and the pharynx breaks down in the fourth week.
The stomodeum is bounded below by the mandibular prominence of the first arch (Fig. 1.25), which produces the floor of the mouth, lower jaw and lower lip. From mesenchyme (embryonic mesoderm) on the ventral surface of the developing brain the frontonasal prominence grows down towards the stomodeum. This is indented by two nasal placodes which develop into nasal pits. These are bounded by medial and lateral nasal prominences that unite to encircle the nostril. From the cranial aspect of the dorsal region of each mandibular prominence, the maxillary prominence grows ventrally above the stomodeum, forming the floor of the orbit and fusing with the lateral nasal prominence along the line of the nasolacrimal duct. The medial nasal prominences merge to form the intermaxillary segment from which develops the philtrum of the upper lip, the part of the upper jaw that carries the four incisor teeth and the adjacent primary palate. The maxillary prominences fuse with the philtrum to form the whole of the upper lip.
|
|
|
Figure 1.25 Stages in the development of the face. |
At first the developing tongue lies against the floor of the cranium. A midline flange (the nasal septum) grows down from the base of the mesenchymal precursor of the skull. From each maxillary process a flange, known as the palatal shelf, grows downwards and medially; these shelves are soon elevated to a horizontal position over the dorsum of the tongue. The two palatal shelves meet and unite, forming the secondary palate. They also fuse with the nasal septum and the primary palate, the midline incisive foramen persisting at the site of the latter fusion. These fusions begin anteriorly during the eighth week and extend posteriorly to become complete at the uvula in the tenth week.
The nerve supply of all these structures is derived from the fifth cranial (trigeminal) nerve. The frontonasal prominence and its derivatives are supplied by the ophthalmic division, the maxillary prominence and its derivatives by the maxillary division and the mandibular prominence and its derivatives by the mandibular division.
Defects of development
The most common abnormalities are cleft lip and cleft palate (1 in 1000 and 1 in 2500 births respectively) and there are ethnic variations in these incidences; they may or may not coexist. Cleft lip is more frequently lateral. The cleft runs down from the nostril and results from a failure of fusion between the maxillary and medial nasal prominences. Cleft lip may be bilateral and may involve the upper jaw and extend between the primary and secondary palates, the central part being an isolated intermaxillary segment.
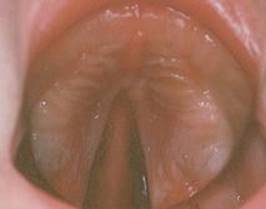
Cleft palate may be partial or complete. As the two palatal processes unite with each other progressively from front to back, arrest of union results in a posterior defect (Fig. 1.26) that varies from the mildest form of bifid uvula to a complete cleft from uvula to gum. In the latter case the cleft almost always runs between a lateral incisor and a canine tooth. Very rarely a midline cleft may separate the two halves of the maxilla. Irregular formations of incisor and canine teeth often accompany these defects of palatal development.
|
|
|
Figure 1.26 Cleft palate. Palatal fusion has commenced anteriorly but a large posterior defect persists. (Provided by Mr S van Eeden, Alder Hey Hospital, Liverpool.) |
A less common defect arises from the failure of fusion of the lateral nasal process with the maxillary process, producing a groove (facial cleft) on the face along the line of the nasolacrimal duct.
Development of the cloaca
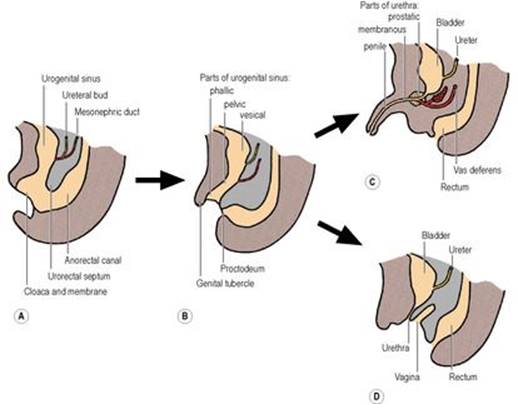
At the caudal end of the embryo, the hindgut and the allantois (a diverticulum from the yolk sac) meet in a common cavity, the cloaca, bounded distally by the cloacal membrane (Fig. 1.27A). From the dorsal wall of the allantois, the urorectal septum grows downwards to meet the cloacal membrane, so dividing the cloaca and membrane into two (Fig. 1.27B): at the front are the urogenital sinus and urogenital membrane, and at the back the anorectal canal and the anal membrane, which lies in a small ectodermal depression, the proctodeum. The tip of the urorectal septum forms the perineal body.
|
|
|
Figure 1.27 Development of the cloaca: A the urorectal septum grows down to divide the cloaca into the urogenital sinus and the anorectal canal; B the uppermost (vesicourethral) part of the sinus becomes the bladder and the proximal part of the prostatic urethra, with the pelvic and phallic parts distally; C in the male the pelvic part becomes the prostatic urethra distal to the opening of the ejaculatory ducts, and the phallic part becomes the dorsal part of the penile urethra; D in the female the bladder and urethra are from the vesicourethral part of the sinus. |
The urogenital sinus (endoderm) has three unequally sized parts. The uppermost and largest is the vesical (vesicourethral) part, which forms most of the bladder epithelium (with surrounding mesoderm forming the muscle and connective tissue) and the female urethra (Fig. 1.27D). The lower end of the mesonephric duct (see p. 286) opens into this part of the sinus, with the ureter arising as a bud from the duct. The lower ends of the duct and ureter become incorporated into the developing bladder, so forming the trigone and in the male the part of the urethra proximal to the opening of the ejaculatory duct (Fig. 1.27C).
The middle or pelvic part of the sinus forms the rest of the prostatic urethra, the membranous urethra and the prostate (with surrounding mesoderm forming the fibromuscular stroma). In the female it contributes to the vagina (derived principally from the paramesonephric ducts; see p. 307).
The lowest or phallic part of the sinus becomes the dorsal part of the penis and penile urethra or the lower part of the vagina. At the front of the urogenital membrane (which breaks down) is a midline mesodermal swelling, the genital tubercle (Fig. 1.27B), which becomes the glans penis or clitoris. Leading back from the tubercle on either side are the urogenital folds, which in the female remain separate as the labia minora. In the male they unite at the back to form the midline raphe of the scrotum, the rest of the scrotum coming from the pair of genital (labioscrotal) swellings which develop lateral to the urogenital folds and which in the female become the labia majora. The front parts of the urogenital folds unite from the scrotum forwards as the ventral part of the penis and penile urethra; failure of such fusion results in hypospadias, where the urethra opens on the ventral aspect of a malformed penis.
Cardiac and venous development
Early development of the heart
Primitive blood vessels are laid down by angioblasts on the wall of the yolk sac. Two such vessels fuse together to make a single heart tube which develops muscle fibres in its wall and becomes pulsatile. It differentiates into four parts which in a cephalocaudal direction are the bulb, ventricle, atrium and sinus venosus. The tube grows at a greater rate than the cavity (the primitive pericardial cavity) in which it is suspended; it therefore has to bend, and it does so in such a way that the bulb and the ventricle come to lie in front of the atrium and sinus venosus. There is also a slight twisting of the bulb to the right and the larger ventricle to the left, hence the normal left-sided bulging of the definitive heart.
The upper part of the bulb is the truncus arteriosus, which divides to become the aorta and pulmonary trunk. The lower part of the bulb becomes most of the right ventricle, with the original ventricle forming most of the left ventricle. The atrium becomes divided into two, with the sinus venosus becoming mostly absorbed into the right atrium.
Development of veins
From a network of primitive veins, certain longitudinal channels develop to return blood to the sinus venosus. It receives blood from three sources: from the placenta by umbilical veins, from the yolk sac (which becomes the alimentary canal) by vitelline veins, and from the general tissues of the embryo by cardinal veins. In each group there are right and left veins, with anastomosing cross-channels between each pair, and the whole or part of one longitudinal vein of each pair disappears: right umbilical; left vitelline; and left cardinal.
The vitelline veins, with their cross-channels, contribute to the formation of the portal vein and the upper end of the inferior vena cava.
The left umbilical vein joins the left branch of the portal vein, but its blood short-circuits the liver by passing along a venous shunt, the ductus venosus, which joins the inferior vena cava on the cranial side of the liver. After birth the left umbilical vein and its continuation, the ductus venosus, become reduced to fibrous cords, the ligamentum teres and ligamentum venosum.
On each side, a vein from the head and neck (internal jugular) and from the upper limb (subclavian) unite to form the anterior cardinal vein. Similarly, from the lower limb and pelvis, external and internal iliac veins form the posterior cardinal vein, into which drain segmental veins (intercostal and lumbar). The anterior and posterior veins unite to form the short common cardinal vein which opens into the sinus venosus. The essential features of subsequent changes are the obliteration of the major portions of the left anterior and posterior cardinals, with the persistence of a cross-channel at each end of the trunk: the left brachiocephalic vein and the left common iliac vein.
In the thorax the right anterior cardinal vein forms the right brachiocephalic vein and part of the superior vena cava, the rest being derived from the common cardinal. The azygos and hemiazygos veins develop from the right posterior cardinal vein and the supracardinal vein that replaces the left posterior cardinal vein.
In the abdomen other longitudinal channels appear, both medial (subcardinal) and dorsal (supracardinal) to the original posterior cardinal. The end result is the formation of the inferior vena cava and its tributaries from different parts of these vessels and their intercommunications. In the lower abdomen the common iliac veins are behind the corresponding arteries, but higher up the renal veins are in front of the renal arteries, due to their development from dorsal or ventral venous channels.
Fetal circulation
The fetal blood is oxygenated in the placenta not in the lungs. The economy of the fetal circulation is improved by three short-circuiting arrangements, all of which cease to function at the time of birth: the ductus venosus, the foramen ovale and the ductus arteriosus.
Ductus venosus
Oxygenated blood returns from the placenta by the (left) umbilical vein, which joins the left branch of the portal vein in the porta hepatis. This oxygenated blood short-circuits the sinusoids of the liver; it is conveyed directly to the inferior vena cava by the ductus venosus. This channel lies along the inferior surface of the liver, between the attached layers of the lesser omentum. After birth, when blood no longer flows along the thrombosed umbilical vein, the blood in the ductus venosus clots and the ductus venosus becomes converted into a fibrous cord, the ligamentum venosum, lying deep in the cleft bounding the caudate lobe of the liver. The intra-abdominal part of the umbilical vein persists as a fibrous cord, the ligamentum teres. The two are continuous.
Foramen ovale
The interatrial septum of the fetal heart is patent, being perforated by the foramen ovale. Blood brought to the right atrium by the inferior vena cava is directed by its ‘valve’ through the foramen and so enters the left atrium. The oxygenated placental blood is thus made to bypass the right ventricle and the airless lungs, and is directed into the left ventricle and aorta and so to the carotid arteries.
After birth the foramen ovale is closed by fusion of the primary and secondary septa (see p. 206). After closure all the blood in the right atrium passes into the right ventricle and so to the lungs.
Ductus arteriosus
It has already been noted that oxygenated blood in the umbilical vein passes via the ductus venosus, inferior vena cava and right atrium through the foramen ovale to the left side of the heart and so to the head. Venous blood from the head is returned by way of the brachiocephalic veins to the superior vena cava. In the right atrium this venous bloodstream crosses the stream of oxygenated blood brought there via the inferior vena cava. The two streams of blood scarcely mix with each other. The deoxygenated blood from the superior vena cava passes through the right atrium into the right ventricle and so into the pulmonary trunk. It now short-circuits the airless lungs by the ductus arteriosus. This is a thick-walled artery joining the left branch of the pulmonary trunk to the aorta, distal to the origin of the three branches of the aortic arch. The deoxygenated blood thus passes distally along the aorta and the common and internal iliac arteries, and via the umbilical arteries, to the placenta to be reoxygenated.
After birth the ductus arteriosus is occluded by contraction of its muscular walls. It persists as a fibrous band, the ligamentum arteriosum, which connects the commencement of the left pulmonary artery to the concavity of the arch of the aorta. The umbilical arteries close off and become fibrous cords: the medial umbilical ligaments.